Potrebbero piacerti anche
- Taller de EspectrometriaDocumento2 pagineTaller de EspectrometriaAnderson Alvarado0% (1)
- AGA Disturbios Respiratorios PLUS MEDIC ADocumento8 pagineAGA Disturbios Respiratorios PLUS MEDIC AGretchen LavadoNessuna valutazione finora
- Farmaco AlopurinolDocumento3 pagineFarmaco AlopurinolJesús Leyva GarcíaNessuna valutazione finora
- Practica 6 Preparacion de PiruvatoDocumento2 paginePractica 6 Preparacion de PiruvatoStefany LugoNessuna valutazione finora
- Practica 13Documento2 paginePractica 13Andre HaroNessuna valutazione finora
- Informe Practica 17 CUERPOS CETÓNICOS EN ORINADocumento7 pagineInforme Practica 17 CUERPOS CETÓNICOS EN ORINALin Arroyo50% (2)
- Acumulos Intracelulares ActualDocumento23 pagineAcumulos Intracelulares ActualMailyn Amparo Gutierrez Delgado50% (2)
- Problemas EnzimasDocumento5 pagineProblemas EnzimasKevin O. BallardoNessuna valutazione finora
- Determinacion de Acido Úrico y de UreaDocumento8 pagineDeterminacion de Acido Úrico y de UreaMIGUELDARIONessuna valutazione finora
- Practica de Acido UricoDocumento13 paginePractica de Acido UricoLuis AlfredoNessuna valutazione finora
- PRACTICA 03b. DEMOSTRACION DE LA ACCIÓN DE LA LIPASA PANCREÁTICA Y SALES BILIARESDocumento2 paginePRACTICA 03b. DEMOSTRACION DE LA ACCIÓN DE LA LIPASA PANCREÁTICA Y SALES BILIARESMaricielo Alva100% (1)
- 10A Metabolismo de La Glucosa, GlucólisisDocumento7 pagine10A Metabolismo de La Glucosa, Glucólisissarita esther100% (1)
- Lab 02. Potenciales de Acción NeuralDocumento16 pagineLab 02. Potenciales de Acción NeuralJUSEYBI CORTEZNessuna valutazione finora
- Actividad 4 y 5 Lab Fisio RanaDocumento3 pagineActividad 4 y 5 Lab Fisio RanaCarlos Jair Herrera CamposNessuna valutazione finora
- 1.introduccion Al MetabolismoDocumento9 pagine1.introduccion Al MetabolismoAndres Felipe Prieto AlarconNessuna valutazione finora
- Practica 1 Bioquimica MetabolicaDocumento5 paginePractica 1 Bioquimica MetabolicaYunhuel PerezNessuna valutazione finora
- Determinación de Acido Urico en SangreDocumento6 pagineDeterminación de Acido Urico en SangreOmar Tostado100% (1)
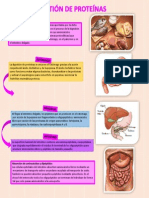
- Digestión de PreteínasDocumento2 pagineDigestión de PreteínasbriandaaliciaanguloNessuna valutazione finora
- PEPTIDASAS PresentacionDocumento15 paginePEPTIDASAS PresentacionFabi MontoyaNessuna valutazione finora
- Práctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Documento11 paginePráctica No.12 "Factores Que Afectan La Velocidad de Reacción Enzimática "Angel Gustavo HernandezNessuna valutazione finora
- Lab. Bioqui Semana 10Documento5 pagineLab. Bioqui Semana 10Aylin ArellanoNessuna valutazione finora
- Determinación de Proteinas Totales en SueroDocumento9 pagineDeterminación de Proteinas Totales en SueroMario LoaizaNessuna valutazione finora
- Informe Efecto de La Concentracion Del Sustracto Calcular KMDocumento8 pagineInforme Efecto de La Concentracion Del Sustracto Calcular KMEstrella OrozcoNessuna valutazione finora
- Bioquimica Ii 1er Parcial OficialDocumento122 pagineBioquimica Ii 1er Parcial OficialAna Gabriela Fernandez Torrez100% (1)
- Determinación de PH y Soluciones AmortiguadorasDocumento3 pagineDeterminación de PH y Soluciones AmortiguadorasBrayan MejíaNessuna valutazione finora
- Guía FarmacoterapeuticaDocumento16 pagineGuía FarmacoterapeuticaAna Laura Ayala100% (2)
- Practica # 6 - Fosfatasa AlcalinaDocumento7 paginePractica # 6 - Fosfatasa Alcalinarockakicha100% (1)
- Amilasa SericaDocumento19 pagineAmilasa Sericayenso2Nessuna valutazione finora
- Targets Mínimos Práctica FisiologíaDocumento3 pagineTargets Mínimos Práctica FisiologíaHady Quispe LeónNessuna valutazione finora
- Amortiguadores FisiologicosDocumento42 pagineAmortiguadores FisiologicosZuffelyEstelaRojas100% (1)
- Informe Determinacion de Urea PDFDocumento7 pagineInforme Determinacion de Urea PDFAna Lucía VillaverdeNessuna valutazione finora
- BANCO DE PREGUNTAS SEGUNDO PARCIAL WiiiiiiiiiiDocumento10 pagineBANCO DE PREGUNTAS SEGUNDO PARCIAL WiiiiiiiiiiMelendez Quispe William JoseNessuna valutazione finora
- Descarboxilacion OxidativaDocumento34 pagineDescarboxilacion Oxidativaelvismartha50% (2)
- Informe 5 Reconocimiento de Vitaminas Helber Rojas, Danny Bautista.Documento10 pagineInforme 5 Reconocimiento de Vitaminas Helber Rojas, Danny Bautista.Helber Fernando Rojas BarajasNessuna valutazione finora
- Determinación de La Actividad Enzimatica de La Amilasa SalivalDocumento12 pagineDeterminación de La Actividad Enzimatica de La Amilasa Salivalclubdelquimico.blogspot.com81% (21)
- PRACTICA 01. Determinación de Ácido ÚricoDocumento3 paginePRACTICA 01. Determinación de Ácido ÚricosilviaNessuna valutazione finora
- Práctica 9. Identtificación de CandidaDocumento21 paginePráctica 9. Identtificación de CandidaSofía Ochoa Astudillo100% (2)
- Monografia Fisiologia HematicaDocumento22 pagineMonografia Fisiologia HematicaKevin SoaresNessuna valutazione finora
- Guia Paso Químico de OrinaDocumento6 pagineGuia Paso Químico de OrinaElizabeth100% (1)
- Práctica 13Documento4 paginePráctica 13Alondra FortunatNessuna valutazione finora
- HHOÍDJCCIFVDocumento15 pagineHHOÍDJCCIFVthais pizarro contreras50% (2)
- Marco TeóricoDocumento8 pagineMarco TeóricoDaniel Arroyo AedoNessuna valutazione finora
- 3 Semana - Practica - Aminoacidos y Proteinas Como Electrolitos - PresentacionDocumento14 pagine3 Semana - Practica - Aminoacidos y Proteinas Como Electrolitos - PresentacionAntonyNessuna valutazione finora
- Resumen Introducción A La Química Clínica y EspectrofotometríaDocumento5 pagineResumen Introducción A La Química Clínica y EspectrofotometríajeiselllaNessuna valutazione finora
- ANON-Apuntes y Lecciones de Química Farmaceútica, Universidad de AlcaláDocumento133 pagineANON-Apuntes y Lecciones de Química Farmaceútica, Universidad de AlcaláJuanManuelAmaroLuisNessuna valutazione finora
- Actividad 02, Sesión Práctica 02Documento4 pagineActividad 02, Sesión Práctica 02Hany MontanoNessuna valutazione finora
- Inhibicion de Cadena Transportadora de ElectronesDocumento4 pagineInhibicion de Cadena Transportadora de ElectronesAaron Llerena ArroyoNessuna valutazione finora
- Sesión 4 Liquidos CorporalesDocumento16 pagineSesión 4 Liquidos CorporalesAnibal Alberto Sosa EscobarNessuna valutazione finora
- Determinacion de Urea CuestionarioDocumento2 pagineDeterminacion de Urea CuestionarioFernando LiviasNessuna valutazione finora
- Actividad 1 Cuestionario 1 de BioquimicaDocumento10 pagineActividad 1 Cuestionario 1 de BioquimicaJairo TavaresNessuna valutazione finora
- Práctica 11 EnzimasDocumento6 paginePráctica 11 EnzimasJeffe Yepes GarcíaNessuna valutazione finora
- Informe CKDocumento4 pagineInforme CKKarla Orozco0% (1)
- Guia de Ejercicios Sobre PHDocumento5 pagineGuia de Ejercicios Sobre PHbarlosNessuna valutazione finora
- Importancia de La Fisicoquímica en La Bioquímica y FarmaciaDocumento4 pagineImportancia de La Fisicoquímica en La Bioquímica y FarmaciaNancyGonzalezNessuna valutazione finora
- Metabolismo de EicosanoidesDocumento16 pagineMetabolismo de EicosanoidesGino Chávez SánchezNessuna valutazione finora
- Reguladores en La SangreDocumento6 pagineReguladores en La SangreJessica Munoz100% (1)
- Inhibidores de La GlucolisisDocumento9 pagineInhibidores de La Glucolisisingjuferdima2240100% (1)
- Monooxigenasas CP 450 Puto IsidroDocumento5 pagineMonooxigenasas CP 450 Puto IsidroMarlon GuerreroNessuna valutazione finora
- QuimicaDocumento6 pagineQuimicaAyelén SanchezNessuna valutazione finora
- Diapositivas Biologia MitocondriaDocumento19 pagineDiapositivas Biologia MitocondriaDanny JosuéNessuna valutazione finora
- Arte y CulturaDocumento7 pagineArte y Culturacarlos stiven vasquez cochachinNessuna valutazione finora
- Hunter X Hunter - Tomo 24 - Absorbiendo MangasDocumento199 pagineHunter X Hunter - Tomo 24 - Absorbiendo Mangascarlos stiven vasquez cochachinNessuna valutazione finora
- TrabajoDocumento1 paginaTrabajocarlos stiven vasquez cochachinNessuna valutazione finora
- Hunter X Hunter - Tomo 16 - Absorbiendo MangasDocumento186 pagineHunter X Hunter - Tomo 16 - Absorbiendo Mangascarlos stiven vasquez cochachinNessuna valutazione finora
- Hunter X Hunter - Tomo 15 - Absorbiendo MangasDocumento191 pagineHunter X Hunter - Tomo 15 - Absorbiendo Mangascarlos stiven vasquez cochachinNessuna valutazione finora
- Hunter X Hunter - Tomo 14 - Absorbiendo MangasDocumento196 pagineHunter X Hunter - Tomo 14 - Absorbiendo Mangascarlos stiven vasquez cochachinNessuna valutazione finora
- Hunter X Hunter - Tomo 13 - Absorbiendo MangasDocumento189 pagineHunter X Hunter - Tomo 13 - Absorbiendo Mangascarlos stiven vasquez cochachinNessuna valutazione finora
- Marco Teorico TDocumento16 pagineMarco Teorico Tcarlos stiven vasquez cochachinNessuna valutazione finora
- Parque EcologicoDocumento18 pagineParque Ecologicocarlos stiven vasquez cochachinNessuna valutazione finora
- AGARDocumento8 pagineAGARcarlos stiven vasquez cochachinNessuna valutazione finora
- LogicaDocumento5 pagineLogicacarlos stiven vasquez cochachin67% (3)
- Unidad N.º 2: Bases Químicas de La VidaDocumento31 pagineUnidad N.º 2: Bases Químicas de La VidaAna Fatecha CabreraNessuna valutazione finora
- Elaboración y Evaluación Nutritiva de Helado A Base de QuinuaDocumento7 pagineElaboración y Evaluación Nutritiva de Helado A Base de Quinuaolger frisancho chuquicallataNessuna valutazione finora
- Informe Proteinas JorgeDocumento20 pagineInforme Proteinas JorgeJ Mora GañanNessuna valutazione finora
- Extraccion de CaseinaDocumento4 pagineExtraccion de CaseinaANiita GAbiiela ArriiagaNessuna valutazione finora
- 03 Agi 209 Articulo CientificoDocumento10 pagine03 Agi 209 Articulo CientificoFreddy Quispe TuncarNessuna valutazione finora
- Trabajo Vidal AscallaDocumento119 pagineTrabajo Vidal AscallaPrudencio PatriciaNessuna valutazione finora
- Indicadores Productivos en GanaderiaDocumento20 pagineIndicadores Productivos en GanaderiaRuben Sebastian Saabedra MarinNessuna valutazione finora
- 59-Paja Tratada Con UreaDocumento7 pagine59-Paja Tratada Con UreaCoral Cruz MadridNessuna valutazione finora
- Cartas Tecnologicas Grupo 25Documento51 pagineCartas Tecnologicas Grupo 25alexanderNessuna valutazione finora
- Enzimas en Los Alimentos PDFDocumento49 pagineEnzimas en Los Alimentos PDFClaudia Daiana Armella Aparicio100% (1)
- Metabolismo de ProteínasDocumento9 pagineMetabolismo de ProteínasGermain EduardOoNessuna valutazione finora
- Requerimientos Nutricionales Del Ganado de Leche Según El Modelo Del NRC PDFDocumento7 pagineRequerimientos Nutricionales Del Ganado de Leche Según El Modelo Del NRC PDFCRISTIAN JOSE GOMEZ ROJASNessuna valutazione finora
- Aplicaciones Foliares de Urea en Árboles FrutalesDocumento2 pagineAplicaciones Foliares de Urea en Árboles Frutalespin_2863Nessuna valutazione finora
- Analítica SeparataDocumento72 pagineAnalítica SeparataHans CáceresNessuna valutazione finora
- Fisiologia Humana Unidad 1Documento8 pagineFisiologia Humana Unidad 1yeridotelNessuna valutazione finora
- Tema 5. Genética Molecular (Biología 4º ESO)Documento7 pagineTema 5. Genética Molecular (Biología 4º ESO)LuNessuna valutazione finora
- Enzimologa ClnicaDocumento80 pagineEnzimologa ClnicaWillian René Rodríguez VázquezNessuna valutazione finora
- Evolución QuímicaDocumento45 pagineEvolución QuímicaNiriaNessuna valutazione finora
- Universidad Ola Habana PDFDocumento84 pagineUniversidad Ola Habana PDFMagaly BenaventeNessuna valutazione finora
- DengueDocumento43 pagineDenguebenito romeroNessuna valutazione finora
- Generalidades Sobre Nutrición y Alimentación de RumiantesDocumento6 pagineGeneralidades Sobre Nutrición y Alimentación de RumiantesJuliana GonzálezNessuna valutazione finora
- Silabo-Quimica Organica (2021-B) DufaDocumento10 pagineSilabo-Quimica Organica (2021-B) DufaKaterin Melipsa Moscoso Esquivel100% (1)
- BiomoléculasDocumento4 pagineBiomoléculasKeWaTeImportaNessuna valutazione finora
- Alimentos para Vacas LecherasDocumento30 pagineAlimentos para Vacas Lecherasingridcordoba9550% (2)
- Mercado - Chocolate AlgarroboDocumento30 pagineMercado - Chocolate AlgarrobojeassonmcraeNessuna valutazione finora
- Identificacion de Aminoacido y ProteinaDocumento3 pagineIdentificacion de Aminoacido y ProteinaMarcos Nuñez FernandezNessuna valutazione finora
- Impacto de La Biotecnologia en El Sector Lacteo Potenciales Nichos de Aplicacion en ChileDocumento25 pagineImpacto de La Biotecnologia en El Sector Lacteo Potenciales Nichos de Aplicacion en ChileElenaNessuna valutazione finora
- Banco de Pregunta de BiologíaDocumento15 pagineBanco de Pregunta de BiologíaMilton Taco AlbitesNessuna valutazione finora
- Purificacion en Power PointDocumento13 paginePurificacion en Power PointkikeburguerNessuna valutazione finora
- Menú Semanal para Seguir El Programa Whole30Documento28 pagineMenú Semanal para Seguir El Programa Whole30Anonymous ToiJi4HqNessuna valutazione finora