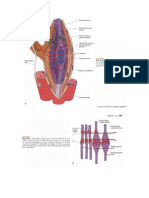
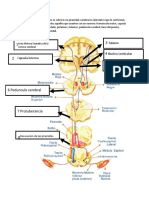
Potrebbero piacerti anche
- MicroencefaliaDocumento12 pagineMicroencefaliaHelen Monsalves EspinozaNessuna valutazione finora
- El Nucleo CelularDocumento4 pagineEl Nucleo CelularVale Constanza100% (1)
- Zonas de Oclusión y ParedDocumento5 pagineZonas de Oclusión y ParedRock Velazco0% (1)
- Huso NeuromuscularDocumento4 pagineHuso NeuromuscularAdriana Cárdenas Guijarro100% (1)
- La Respiración Aeróbica y La MitocondriaDocumento2 pagineLa Respiración Aeróbica y La MitocondriaKarla Nicole Silverio TaverasNessuna valutazione finora
- Habilidades Motrices BásicasDocumento6 pagineHabilidades Motrices BásicasJoseffaArancibiaNessuna valutazione finora
- Médula EspinalDocumento9 pagineMédula EspinalKarina GuevaraNessuna valutazione finora
- Arco Reflejo LibroDocumento109 pagineArco Reflejo LibroMarlen AcostaNessuna valutazione finora
- Proceso de Mielinización o MielyogenesisDocumento5 pagineProceso de Mielinización o MielyogenesisEugenia Carabajal0% (1)
- Sistema Nervioso CentralDocumento7 pagineSistema Nervioso CentralRoger lima ponce0% (1)
- Tema 5. To Basada en La EvidenciaDocumento7 pagineTema 5. To Basada en La EvidenciaNatalia Lucena MenaNessuna valutazione finora
- Reflejo OsteotendinosoDocumento17 pagineReflejo OsteotendinosoJessica León67% (3)
- VÍAS MOTORAS y SENSITIVASDocumento5 pagineVÍAS MOTORAS y SENSITIVASSabrina Rivero100% (5)
- Tejido Conectivo o Conjuntivo PDFDocumento18 pagineTejido Conectivo o Conjuntivo PDFLuis SingerNessuna valutazione finora
- Ensayo Medios FisicosDocumento7 pagineEnsayo Medios FisicosvictorNessuna valutazione finora
- Piramidal y Extrapiramidal - BuenoDocumento7 paginePiramidal y Extrapiramidal - BuenodtNessuna valutazione finora
- Resumen Tejido Conjuntivo o ConectivoDocumento7 pagineResumen Tejido Conjuntivo o ConectivoOrlandoArtemioPoloVeraNessuna valutazione finora
- Tejido Conjuntivo-InfografiaDocumento1 paginaTejido Conjuntivo-InfografiaAmérica MarceloNessuna valutazione finora
- Practica Exploración de ReflejosDocumento9 paginePractica Exploración de ReflejosNadia MoraNessuna valutazione finora
- Leccion06. Embriologia Del Aparato Locomotor - Desarrollo Del Sistema EsqueléticoDocumento4 pagineLeccion06. Embriologia Del Aparato Locomotor - Desarrollo Del Sistema EsqueléticoLaura Gozalbo Nieto100% (8)
- Permeabilidad MembranalDocumento5 paginePermeabilidad MembranalANGEL ERNESTO ABARCA RODRIGUEZNessuna valutazione finora
- Caracteristicas Fisicas de La CirculaciónDocumento13 pagineCaracteristicas Fisicas de La CirculaciónJhon Cruz67% (3)
- Historia de La BiofisicaDocumento1 paginaHistoria de La BiofisicaDiana De JesusNessuna valutazione finora
- 01 Tejido MuscularDocumento28 pagine01 Tejido Muscularmanuela castroNessuna valutazione finora
- Mapa Conceptual Sistema EsqueleticoDocumento1 paginaMapa Conceptual Sistema EsqueleticoJhaquelin RodriguezNessuna valutazione finora
- Carta Descriptiva Procesos Biológicos 22-2Documento21 pagineCarta Descriptiva Procesos Biológicos 22-2Ruth Cobo RosalesNessuna valutazione finora
- Tejido Nervioso. HistologiaDocumento10 pagineTejido Nervioso. HistologiaManuel ChervoNessuna valutazione finora
- Diapositivas ExposiciónDocumento33 pagineDiapositivas ExposiciónNataly PillajoNessuna valutazione finora
- Conductividad NerviosaDocumento2 pagineConductividad NerviosaAlexanders Mercado100% (1)
- NeuroplasticidadDocumento1 paginaNeuroplasticidadAna BotelloNessuna valutazione finora
- Cuestionario Nivel CelularDocumento27 pagineCuestionario Nivel CelularDulce MarïaNessuna valutazione finora
- (8 Organización Funcional Del Sistema Motor)Documento12 pagine(8 Organización Funcional Del Sistema Motor)Orlando J SierraNessuna valutazione finora
- Cadenas CinematicasDocumento4 pagineCadenas CinematicasFlorencia MarengoNessuna valutazione finora
- RADIOAUTOGRAFIA - Futura MédicaDocumento9 pagineRADIOAUTOGRAFIA - Futura MédicafuturamedicaNessuna valutazione finora
- NEUROPLASTICIDADDocumento7 pagineNEUROPLASTICIDADRobert LopezNessuna valutazione finora
- Clase V Periodo Embrionario (Tercera - Octava Semana)Documento83 pagineClase V Periodo Embrionario (Tercera - Octava Semana)JorgeAlonsoSaavedraTorresNessuna valutazione finora
- Cadena CineticaDocumento2 pagineCadena CineticaYENNIBETH GIMENEZNessuna valutazione finora
- Reacciones Inmunopatológicas A Las ParasitosisDocumento2 pagineReacciones Inmunopatológicas A Las Parasitosis3SacNessuna valutazione finora
- ARTROLOGIa - PPT DiapoDocumento52 pagineARTROLOGIa - PPT DiapoDiana Tenorio100% (5)
- Reticulo Endoplasmatico Liso y Rugoso, FuncionesDocumento5 pagineReticulo Endoplasmatico Liso y Rugoso, FuncionesAna MarcelaNessuna valutazione finora
- Esquemas Clase EvcDocumento2 pagineEsquemas Clase EvcIván Gustavo Magaña CeballosNessuna valutazione finora
- Palancas AnatómicasDocumento15 paginePalancas AnatómicasLuisGálvezCastilloNessuna valutazione finora
- Vías OlfatoriaDocumento7 pagineVías OlfatoriaNicole SaaNessuna valutazione finora
- Neuro 2 PDFDocumento4 pagineNeuro 2 PDFRamon Vicente Linares PerezNessuna valutazione finora
- Citoplasma y CitosolDocumento7 pagineCitoplasma y CitosolAlmacenCiberPreferidoNessuna valutazione finora
- Cuestionario de Sistema Endocrino, HistologíaDocumento10 pagineCuestionario de Sistema Endocrino, HistologíaDayana IntriagoNessuna valutazione finora
- 6 Musculos Que Mueven El Humero (Hueso DelDocumento22 pagine6 Musculos Que Mueven El Humero (Hueso DelbetoNessuna valutazione finora
- Clasificacion de Fibras NerviosasDocumento49 pagineClasificacion de Fibras NerviosasVictor Black100% (1)
- Guia de Prácticas de Laboratorio (Anatomía)Documento3 pagineGuia de Prácticas de Laboratorio (Anatomía)Mauri GuañunaNessuna valutazione finora
- Linea Del Tiempo Fisioterapia PDFDocumento2 pagineLinea Del Tiempo Fisioterapia PDFStefanii Mendivil RomeroNessuna valutazione finora
- Control Motor 1Documento79 pagineControl Motor 1Matilde Espinoza OlivaresNessuna valutazione finora
- MF 2 - Guía 04 - Sistema LinfáticoDocumento8 pagineMF 2 - Guía 04 - Sistema LinfáticoYasmin RimariNessuna valutazione finora
- Cuestionario Sistema VestibularDocumento1 paginaCuestionario Sistema VestibularRonny Jose Blanco Valdivieso0% (1)
- Funciones Secretoras Del Tubo DigestivoDocumento7 pagineFunciones Secretoras Del Tubo DigestivoSiIvana VCNessuna valutazione finora
- Sistemas de Endomembranas, Resumen Capítulo 8, Gerard Karp Biología Celular y Molecular, 6 EdiciónDocumento7 pagineSistemas de Endomembranas, Resumen Capítulo 8, Gerard Karp Biología Celular y Molecular, 6 Ediciónsuldeny100% (2)
- Glosario de AnatomiaDocumento6 pagineGlosario de AnatomiaGatita MiauNessuna valutazione finora
- Trafico Intracelular-SulyDocumento3 pagineTrafico Intracelular-SulySuly VigoNessuna valutazione finora
- Aves Clase 2 Fisiopatologia 2018Documento86 pagineAves Clase 2 Fisiopatologia 2018Jeannett Yolanda Noa LeonNessuna valutazione finora
- Adherencia CelularDocumento14 pagineAdherencia CelularAndy Paúl García OrbeNessuna valutazione finora
- Mac y MatrizDocumento20 pagineMac y MatrizMarce De La CruzNessuna valutazione finora
- Practica 3 BioquímicaDocumento1 paginaPractica 3 BioquímicaFlor Carrera50% (2)
- GRANULACIÓNDocumento45 pagineGRANULACIÓNFlor Carrera50% (2)
- Anatomía Del Intestino DelgadoDocumento7 pagineAnatomía Del Intestino DelgadoFlor CarreraNessuna valutazione finora
- Prácticas MicrobiologíaDocumento10 paginePrácticas MicrobiologíaFlor CarreraNessuna valutazione finora
- Sistemas Coloidales y Umbral de CoagulaciónDocumento10 pagineSistemas Coloidales y Umbral de CoagulaciónFlor Carrera100% (1)
- Actividad Superficial de Alcoholes AlifaticosDocumento13 pagineActividad Superficial de Alcoholes AlifaticosFlor Carrera67% (3)
- Absorción de I2 Por Ácidos Grasos InsaturadosDocumento3 pagineAbsorción de I2 Por Ácidos Grasos InsaturadosFlor Carrera67% (3)
- Determinación de Agua Con El Reactivo de Karl FischerDocumento3 pagineDeterminación de Agua Con El Reactivo de Karl FischerFlor CarreraNessuna valutazione finora
- Practica No 8. MicrobiologiaDocumento5 paginePractica No 8. MicrobiologiaFlor CarreraNessuna valutazione finora
- ColoidesDocumento34 pagineColoidesFlor Carrera100% (1)
- Tecnología LíticaDocumento24 pagineTecnología LíticaFlor Carrera100% (1)
- Cuestionario de Antigeno ProstaticoDocumento2 pagineCuestionario de Antigeno ProstaticoFlor CarreraNessuna valutazione finora
- ESQUEMAS - Diagrama de Esquemas Maladaptativos TempranosDocumento5 pagineESQUEMAS - Diagrama de Esquemas Maladaptativos TempranosMariella100% (3)
- Contrato NuevoDocumento5 pagineContrato NuevoBayardo GadeaNessuna valutazione finora
- Control de Lectura - El Olvido Que Seremos PDFDocumento4 pagineControl de Lectura - El Olvido Que Seremos PDFAlejandra CalderonNessuna valutazione finora
- GARCIA - MARTA - El Paisaje Del Bienestar - Arquitectura Moderna para El Baño Junto Al Rio ManzanaresDocumento505 pagineGARCIA - MARTA - El Paisaje Del Bienestar - Arquitectura Moderna para El Baño Junto Al Rio ManzanarespablopaezgNessuna valutazione finora
- Ensayo-De Lor Rescursos para La Atención Materno-Infantil - IctDocumento5 pagineEnsayo-De Lor Rescursos para La Atención Materno-Infantil - Ictjose manuel rangel portilloNessuna valutazione finora
- Rol de Turnos Gob Oct 23Documento48 pagineRol de Turnos Gob Oct 23jhessicarojas90Nessuna valutazione finora
- Diseño de Un Biorreactor para La Fabricación de CervezaDocumento24 pagineDiseño de Un Biorreactor para La Fabricación de CervezaLiova Vera veraNessuna valutazione finora
- Catalogo ZitroDocumento33 pagineCatalogo ZitroJuan carlosNessuna valutazione finora
- 3 Nanawa Un Error EstratégicoDocumento13 pagine3 Nanawa Un Error EstratégicoJuan Carlos Marquez SejasNessuna valutazione finora
- Tipos de Anomalías GeoquímicasDocumento8 pagineTipos de Anomalías GeoquímicasGabriela solis vazquezNessuna valutazione finora
- Como Poner Una Tienda de Videojuegos - Guía de NegocioDocumento4 pagineComo Poner Una Tienda de Videojuegos - Guía de NegocioAlbertoNessuna valutazione finora
- RD 079-2022-Lp-DeDocumento9 pagineRD 079-2022-Lp-Decesar simonNessuna valutazione finora
- Deshidratación OsmóticaDocumento14 pagineDeshidratación OsmóticaKerlynNessuna valutazione finora
- Apostila de GregoDocumento105 pagineApostila de GregoPaula AranhaNessuna valutazione finora
- Siembra de Muestras Microbiológicas CASR - AspxDocumento13 pagineSiembra de Muestras Microbiológicas CASR - AspxJaviera Viviana Véliz SilvaNessuna valutazione finora
- Planeamiento Estrategico (5 Ciclo)Documento10 paginePlaneamiento Estrategico (5 Ciclo)CoQa Gu GullianNessuna valutazione finora
- Características de La EducaciónDocumento4 pagineCaracterísticas de La EducaciónLibreria La HuitecaNessuna valutazione finora
- La Numismática en El PerúDocumento26 pagineLa Numismática en El PerúYessenia Ocsa SerranoNessuna valutazione finora
- Ficha para Diagnóstico de Una IE 2016Documento7 pagineFicha para Diagnóstico de Una IE 2016Ci BautistaNessuna valutazione finora
- Tesis Lic Hector David Argueta GranadosDocumento120 pagineTesis Lic Hector David Argueta GranadoswillimasNessuna valutazione finora
- Nte Inen 754 - Prevención de Incendios. Puertas Cortafuego. Requisitos GeneralesDocumento10 pagineNte Inen 754 - Prevención de Incendios. Puertas Cortafuego. Requisitos GeneralesdarioNessuna valutazione finora
- Capitulo 3 - Economía InternacionalDocumento7 pagineCapitulo 3 - Economía InternacionalCRISTIAN CAMILO CIFUENTES RAMIREZNessuna valutazione finora
- Diccionario de DatosDocumento14 pagineDiccionario de DatosWilliam LozadoNessuna valutazione finora
- 01-08-2022 DesmovilizaciónDocumento15 pagine01-08-2022 DesmovilizaciónJoe Eden QNessuna valutazione finora
- Fac 004-002-000000219Documento1 paginaFac 004-002-000000219MOOM SublimacionesNessuna valutazione finora
- Ejemplo Contestacion Demanda de DivorcioDocumento3 pagineEjemplo Contestacion Demanda de DivorcioErnesto Carlos BerrocalNessuna valutazione finora
- Pruebas de Conocimiento y PsicotecnicasDocumento13 paginePruebas de Conocimiento y PsicotecnicasAlejandro Posada SaninNessuna valutazione finora
- Bim Art 3 IV PeriodoDocumento2 pagineBim Art 3 IV PeriodoisabelNessuna valutazione finora
- Problemas Perceptuales y SensorialesDocumento14 pagineProblemas Perceptuales y SensorialesDanny Pimentel100% (1)
- Contestacion Demanda Patricia LelalDocumento12 pagineContestacion Demanda Patricia LelalCarlos Javier Dorado PerafanNessuna valutazione finora