Potrebbero piacerti anche
- Interferon AlphaDocumento22 pagineInterferon AlphaFedri BaysarNessuna valutazione finora
- Interferon AlphaDocumento22 pagineInterferon AlphaFedri BaysarNessuna valutazione finora
- Interferon Gamma An Overview of Signals, Mechanisms and FunctionsDocumento27 pagineInterferon Gamma An Overview of Signals, Mechanisms and FunctionsDimas ErlanggaNessuna valutazione finora
- Classification of Interferons (Ifns) : Interferons and Chemokines Cytokine Receptors/Signaling Adhesion MoleculesDocumento10 pagineClassification of Interferons (Ifns) : Interferons and Chemokines Cytokine Receptors/Signaling Adhesion MoleculesLucas AresNessuna valutazione finora
- InterferonDocumento9 pagineInterferonParabesh DeNessuna valutazione finora
- 2008 - Interferons and Viruses - An Interplay Betweeninduction, Signalling, Antiviral Responses and ViruscountermeasuresDocumento47 pagine2008 - Interferons and Viruses - An Interplay Betweeninduction, Signalling, Antiviral Responses and ViruscountermeasuresFernando Fernández GarcíaNessuna valutazione finora
- 2004 - Interferon Gamma - An Overview of Signals, Mechanismsand FunctionsDocumento27 pagine2004 - Interferon Gamma - An Overview of Signals, Mechanismsand Functions顏世隆Nessuna valutazione finora
- Dissecting Interferon-Induced Transcriptional Programs in Human Peripheral Blood CellsDocumento13 pagineDissecting Interferon-Induced Transcriptional Programs in Human Peripheral Blood CellsKatyusha21Nessuna valutazione finora
- IFN Gamma2Documento8 pagineIFN Gamma2Muhammad IrawanNessuna valutazione finora
- Biochemistry of InterferonDocumento10 pagineBiochemistry of InterferonRai Abdur RehmanNessuna valutazione finora
- Interferon GammaDocumento27 pagineInterferon GammaMarcos MendezNessuna valutazione finora
- Anti IFN Viru ResearchDocumento8 pagineAnti IFN Viru ResearchMohsan UllahNessuna valutazione finora
- Type I and Type III Interferons - Induction, Signaling, Evasion, and Application To Combat COVID-19Documento9 pagineType I and Type III Interferons - Induction, Signaling, Evasion, and Application To Combat COVID-19sprfyllisNessuna valutazione finora
- Type I Interferons and Their Role in Viral InfectionDocumento32 pagineType I Interferons and Their Role in Viral InfectiongauhamiNessuna valutazione finora
- 2016 - The Unique Role of STAT2 in Constitutive and IFN-Induced Transcription and Antiviral ResponsesDocumento11 pagine2016 - The Unique Role of STAT2 in Constitutive and IFN-Induced Transcription and Antiviral ResponsesFernando Fernández GarcíaNessuna valutazione finora
- ReviewDocumento18 pagineReviewMUHAMMAD FARHABNessuna valutazione finora
- 2010 - The differential activity of interferon-α subtypes is consistent among distinct target genes and cell typesDocumento8 pagine2010 - The differential activity of interferon-α subtypes is consistent among distinct target genes and cell typesFernando Fernández GarcíaNessuna valutazione finora
- Avian VirusesDocumento10 pagineAvian VirusesMohsan UllahNessuna valutazione finora
- 2011 - The Two Faces of Interferon-γ in cancerDocumento12 pagine2011 - The Two Faces of Interferon-γ in cancerFernando Fernández GarcíaNessuna valutazione finora
- 2.imunitas PD FilariasisDocumento20 pagine2.imunitas PD FilariasisZalfa Salsabila ApriliaNessuna valutazione finora
- Reviews: RIG-I - Like Receptors: Their Regulation and Roles in RNA SensingDocumento15 pagineReviews: RIG-I - Like Receptors: Their Regulation and Roles in RNA SensingAllan Depieri CataneoNessuna valutazione finora
- 2012 - Activation and Regulation of Interferon-B in Immune ResponsesDocumento16 pagine2012 - Activation and Regulation of Interferon-B in Immune ResponsesFernando Fernández GarcíaNessuna valutazione finora
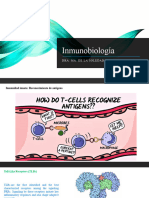
- InmunobiologíaDocumento37 pagineInmunobiologíaSoledad Lagunes CastroNessuna valutazione finora
- 2004 - Characterization of The Murine Alpha Interferon Gene FamilyDocumento10 pagine2004 - Characterization of The Murine Alpha Interferon Gene FamilyFernando Fernández GarcíaNessuna valutazione finora
- Type I Interferon: Friend or Foe?: ReviewDocumento11 pagineType I Interferon: Friend or Foe?: ReviewKraken UrNessuna valutazione finora
- 2010 - Intracellular Expression of IRF9 Stat Fusion Protein Overcomes The Defective Jak-Stat Signaling and Inhibits HCV RNA ReplicationDocumento12 pagine2010 - Intracellular Expression of IRF9 Stat Fusion Protein Overcomes The Defective Jak-Stat Signaling and Inhibits HCV RNA ReplicationFernando Fernández GarcíaNessuna valutazione finora
- Savarin 2008Documento8 pagineSavarin 2008IskvcNessuna valutazione finora
- InterferonDocumento6 pagineInterferonMonika SharmaNessuna valutazione finora
- Interferons: Prepared by MELIS BECERENDocumento14 pagineInterferons: Prepared by MELIS BECERENderyaaaNessuna valutazione finora
- tmp7301 TMPDocumento4 paginetmp7301 TMPFrontiersNessuna valutazione finora
- 58 19Documento10 pagine58 19Alberto RamirezNessuna valutazione finora
- Gof Stat 1Documento31 pagineGof Stat 1Claudia Ivette Aguilar RamosNessuna valutazione finora
- Interferon Genes-1Documento20 pagineInterferon Genes-1Jun KooNessuna valutazione finora
- IRF5 Promotes Inflammatory Macrophage Polarization and T 1-T 17 ResponsesDocumento9 pagineIRF5 Promotes Inflammatory Macrophage Polarization and T 1-T 17 ResponsesMar AlexNessuna valutazione finora
- Natural Killer Cells and Innate Immunity To Protozoan PathogensDocumento12 pagineNatural Killer Cells and Innate Immunity To Protozoan PathogensthehayNessuna valutazione finora
- Briefing DocumentDocumento6 pagineBriefing DocumentHuzaifa KhanNessuna valutazione finora
- Opal2000 PDFDocumento11 pagineOpal2000 PDFKrisztián TóthNessuna valutazione finora
- RoferonDocumento11 pagineRoferonisnaini fahru raziNessuna valutazione finora
- Nishikawa Et Al 2001 in The Absence of Endogenous Gamma Interferon Mice Acutely Infected With Neospora Caninum SuccumbDocumento7 pagineNishikawa Et Al 2001 in The Absence of Endogenous Gamma Interferon Mice Acutely Infected With Neospora Caninum Succumbrifkyfauzan725Nessuna valutazione finora
- Differential Regulation of Human Papillomavirus Type 8 by Interferon Regulatory Factors 3 and 7Documento11 pagineDifferential Regulation of Human Papillomavirus Type 8 by Interferon Regulatory Factors 3 and 7Cahyo GuntoroNessuna valutazione finora
- C04 Innate ImmunityDocumento31 pagineC04 Innate ImmunityforeveraldyNessuna valutazione finora
- ReadingGuide Ch.33 (Acheson)Documento6 pagineReadingGuide Ch.33 (Acheson)Alex CarricoNessuna valutazione finora
- Submitted By: Lovepreet Kaur UID - 14BBT1054Documento20 pagineSubmitted By: Lovepreet Kaur UID - 14BBT1054Anonymous D5g37JjpGBNessuna valutazione finora
- Mora 2006Documento8 pagineMora 2006kevin gelaudeNessuna valutazione finora
- The Role of Interleukin-1 in General PathologyDocumento16 pagineThe Role of Interleukin-1 in General Pathologybaotrann.dangngocNessuna valutazione finora
- Nihms 1573480Documento26 pagineNihms 1573480bilou hertNessuna valutazione finora
- Anti Viral Pathways Innate ImmunityDocumento7 pagineAnti Viral Pathways Innate ImmunityMateus PedrottiNessuna valutazione finora
- Molecular Characterization and Functional Analysis of IRF3 in Tilapia (Oreochromis Niloticus)Documento8 pagineMolecular Characterization and Functional Analysis of IRF3 in Tilapia (Oreochromis Niloticus)Rizka Rahmana PutriNessuna valutazione finora
- Download Genome Wide Identification And Evolution Of Interleukins And Their Potential Roles In Response To Gcrv And Aeromonas Hydrophila Challenge In Grass Carp Ctenopharyngodon Idella Tianbing Xu Zhensh full chapterDocumento42 pagineDownload Genome Wide Identification And Evolution Of Interleukins And Their Potential Roles In Response To Gcrv And Aeromonas Hydrophila Challenge In Grass Carp Ctenopharyngodon Idella Tianbing Xu Zhensh full chapterkevin.walters798100% (4)
- CytokineDocumento7 pagineCytokineMaryam ShahrasbiNessuna valutazione finora
- Lectures 10 (Linked To 12) : Cytokines and Immune ResponseDocumento32 pagineLectures 10 (Linked To 12) : Cytokines and Immune ResponseMac Kevin MandapNessuna valutazione finora
- Antigen Recognition DefrancoDocumento8 pagineAntigen Recognition DefrancoDr.Santosh KumarNessuna valutazione finora
- Toward Targeting in Ammasomes: Insights Into Their Regulation and ActivationDocumento13 pagineToward Targeting in Ammasomes: Insights Into Their Regulation and ActivationRin ChanNessuna valutazione finora
- Economy of Pakistan: Assignment OnDocumento5 pagineEconomy of Pakistan: Assignment OnArman AliNessuna valutazione finora
- Immunology Uworld Notes (Step 1)Documento12 pagineImmunology Uworld Notes (Step 1)Burkitt's LymphomaNessuna valutazione finora
- Gazzinelli 1996Documento13 pagineGazzinelli 1996Tran Trang AnhNessuna valutazione finora
- TNF Superfamily and ILC2 Activation in Asthma by Takahiro Matsuyama, Brittany Marie Salter, Nahal Emami Fard, Kentaro Machida Androma SehmiDocumento19 pagineTNF Superfamily and ILC2 Activation in Asthma by Takahiro Matsuyama, Brittany Marie Salter, Nahal Emami Fard, Kentaro Machida Androma SehmiTedNessuna valutazione finora
- Flu 1Documento10 pagineFlu 1Mau17darwinNessuna valutazione finora
- 2006 - Type I Inteferon Gene Inductionby The Interferon Regulatory FactorFamily of Transcription FactorsDocumento12 pagine2006 - Type I Inteferon Gene Inductionby The Interferon Regulatory FactorFamily of Transcription FactorsFernando Fernández GarcíaNessuna valutazione finora
- Lymphokines: A Forum for Immunoregulatory Cell ProductsDa EverandLymphokines: A Forum for Immunoregulatory Cell ProductsEdgar PickNessuna valutazione finora
- Cover Farmakologi Dasar & Klinik Edisi 10 PDFDocumento1 paginaCover Farmakologi Dasar & Klinik Edisi 10 PDFjidat lawakNessuna valutazione finora
- Western Blotting: DPP4 Enzyme ActivityDocumento2 pagineWestern Blotting: DPP4 Enzyme Activityjidat lawakNessuna valutazione finora
- Embryology: The Anatomic Basis of Fetal MedicineDocumento55 pagineEmbryology: The Anatomic Basis of Fetal Medicinejidat lawakNessuna valutazione finora
- Bucar 2013Documento21 pagineBucar 2013jidat lawakNessuna valutazione finora
- Quetin Leclercq 2004 1Documento27 pagineQuetin Leclercq 2004 1jidat lawakNessuna valutazione finora
- Blackwell S Underground Clinical Vignettes Microbiology PDFDocumento132 pagineBlackwell S Underground Clinical Vignettes Microbiology PDFjidat lawakNessuna valutazione finora
- Tobac AJPM RecsDocumento7 pagineTobac AJPM Recsjidat lawakNessuna valutazione finora
- Allelopathy PDFDocumento16 pagineAllelopathy PDFjidat lawakNessuna valutazione finora
- Rules For SIMPIC 2015 CompetitionDocumento9 pagineRules For SIMPIC 2015 Competitionjidat lawakNessuna valutazione finora
- Rules 2Documento13 pagineRules 2jidat lawakNessuna valutazione finora
- Cable To Metal Surface, Cathodic - CAHAAW3Documento2 pagineCable To Metal Surface, Cathodic - CAHAAW3lhanx2Nessuna valutazione finora
- Lesson 6 - Vibration ControlDocumento62 pagineLesson 6 - Vibration ControlIzzat IkramNessuna valutazione finora
- Head Coverings BookDocumento86 pagineHead Coverings BookRichu RosarioNessuna valutazione finora
- Sakui, K., & Cowie, N. (2012) - The Dark Side of Motivation - Teachers' Perspectives On 'Unmotivation'. ELTJ, 66 (2), 205-213.Documento9 pagineSakui, K., & Cowie, N. (2012) - The Dark Side of Motivation - Teachers' Perspectives On 'Unmotivation'. ELTJ, 66 (2), 205-213.Robert HutchinsonNessuna valutazione finora
- Model 255 Aerosol Generator (Metone)Documento20 pagineModel 255 Aerosol Generator (Metone)Ali RizviNessuna valutazione finora
- FIRST SUMMATIVE EXAMINATION IN ORAL COMMUNICATION IN CONTEXT EditedDocumento3 pagineFIRST SUMMATIVE EXAMINATION IN ORAL COMMUNICATION IN CONTEXT EditedRodylie C. CalimlimNessuna valutazione finora
- Resume: Mr. Shubham Mohan Deokar E-MailDocumento2 pagineResume: Mr. Shubham Mohan Deokar E-MailAdv Ranjit Shedge PatilNessuna valutazione finora
- CV Augusto Brasil Ocampo MedinaDocumento4 pagineCV Augusto Brasil Ocampo MedinaAugusto Brasil Ocampo MedinaNessuna valutazione finora
- EqualLogic Release and Support Policy v25Documento7 pagineEqualLogic Release and Support Policy v25du2efsNessuna valutazione finora
- Mosfet Irfz44Documento8 pagineMosfet Irfz44huynhsang1979Nessuna valutazione finora
- Development Developmental Biology EmbryologyDocumento6 pagineDevelopment Developmental Biology EmbryologyBiju ThomasNessuna valutazione finora
- Close Enough To Touch by Victoria Dahl - Chapter SamplerDocumento23 pagineClose Enough To Touch by Victoria Dahl - Chapter SamplerHarlequinAustraliaNessuna valutazione finora
- 6GK74435DX040XE0 Datasheet enDocumento3 pagine6GK74435DX040XE0 Datasheet enLuis CortezNessuna valutazione finora
- Health Post - Exploring The Intersection of Work and Well-Being - A Guide To Occupational Health PsychologyDocumento3 pagineHealth Post - Exploring The Intersection of Work and Well-Being - A Guide To Occupational Health PsychologyihealthmailboxNessuna valutazione finora
- Determination Rules SAP SDDocumento2 pagineDetermination Rules SAP SDkssumanthNessuna valutazione finora
- In Flight Fuel Management and Declaring MINIMUM MAYDAY FUEL-1.0Documento21 pagineIn Flight Fuel Management and Declaring MINIMUM MAYDAY FUEL-1.0dahiya1988Nessuna valutazione finora
- Week-3-Q1-Gen Chem-Sep-11-15-DllDocumento12 pagineWeek-3-Q1-Gen Chem-Sep-11-15-DllJennette BelliotNessuna valutazione finora
- How To Block HTTP DDoS Attack With Cisco ASA FirewallDocumento4 pagineHow To Block HTTP DDoS Attack With Cisco ASA Firewallabdel taibNessuna valutazione finora
- Eggermont 2019 ABRDocumento15 pagineEggermont 2019 ABRSujeet PathakNessuna valutazione finora
- RFID Seminar AbstractDocumento2 pagineRFID Seminar Abstractanushabhagawath80% (5)
- Heimbach - Keeping Formingfabrics CleanDocumento4 pagineHeimbach - Keeping Formingfabrics CleanTunç TürkNessuna valutazione finora
- Fundaciones Con PilotesDocumento48 pagineFundaciones Con PilotesReddy M.Ch.Nessuna valutazione finora
- Kyle Pape - Between Queer Theory and Native Studies, A Potential For CollaborationDocumento16 pagineKyle Pape - Between Queer Theory and Native Studies, A Potential For CollaborationRafael Alarcón Vidal100% (1)
- Lecture 2 Effects of Operating Conditions in VCCDocumento9 pagineLecture 2 Effects of Operating Conditions in VCCDeniell Joyce MarquezNessuna valutazione finora
- Literatura Tecnica 3Documento10 pagineLiteratura Tecnica 3Christian PerezNessuna valutazione finora
- Activity Title: Learning Targets: Reference (S)Documento5 pagineActivity Title: Learning Targets: Reference (S)Jhev LeopandoNessuna valutazione finora
- LSCM Course OutlineDocumento13 pagineLSCM Course OutlineDeep SachetiNessuna valutazione finora
- UltimateBeginnerHandbookPigeonRacing PDFDocumento21 pagineUltimateBeginnerHandbookPigeonRacing PDFMartinPalmNessuna valutazione finora
- Rom 2 - 0-11 (En)Documento132 pagineRom 2 - 0-11 (En)Mara HerreraNessuna valutazione finora
- Pg2022 ResultDocumento86 paginePg2022 ResultkapilNessuna valutazione finora