Potrebbero piacerti anche
- Temolines. Los coleópteros entre los antiguos mexicanosDa EverandTemolines. Los coleópteros entre los antiguos mexicanosNessuna valutazione finora
- Sinopsis de la Familia Phengodidae (Coleoptera): Trenecitos, bigotudos, glow-worms, railroad-worms o besouros trem de ferroDa EverandSinopsis de la Familia Phengodidae (Coleoptera): Trenecitos, bigotudos, glow-worms, railroad-worms o besouros trem de ferroNessuna valutazione finora
- Salamandra PDFDocumento16 pagineSalamandra PDFanzaeduardo1Nessuna valutazione finora
- Aguila CalzadaDocumento31 pagineAguila Calzadastardust32Nessuna valutazione finora
- Scivul v2Documento14 pagineScivul v2Pintorcito HappinessuwuNessuna valutazione finora
- Lavandera Boyera PDFDocumento11 pagineLavandera Boyera PDFanzaeduardo1Nessuna valutazione finora
- Aegmon v11Documento41 pagineAegmon v11Sara Gómez BallesterosNessuna valutazione finora
- Muserm v2Documento11 pagineMuserm v2Jimmy LoorNessuna valutazione finora
- Aguila AbubillaDocumento33 pagineAguila AbubillagioNessuna valutazione finora
- UrracaDocumento8 pagineUrracaNorma Luz RiveraNessuna valutazione finora
- 5 Atinzaretal Enciclopedia12Documento44 pagine5 Atinzaretal Enciclopedia12Khaled ConstantineNessuna valutazione finora
- Corneja PDFDocumento23 pagineCorneja PDFanzaeduardo1Nessuna valutazione finora
- Lynpar v4Documento33 pagineLynpar v4JpNessuna valutazione finora
- Parmaj v2Documento47 pagineParmaj v2jt892351Nessuna valutazione finora
- Alycis v8Documento19 pagineAlycis v8pilarNessuna valutazione finora
- Tipos OsosDocumento36 pagineTipos Ososalex santafeNessuna valutazione finora
- Rabilargo PDFDocumento17 pagineRabilargo PDFanzaeduardo1Nessuna valutazione finora
- 1 Gorriones PDFDocumento43 pagine1 Gorriones PDFawsedrftgkkswswedNessuna valutazione finora
- Taller AnfibiosDocumento2 pagineTaller AnfibiosLuis Fernando Sáenz0% (1)
- Procyon LotorDocumento31 pagineProcyon LotorCristina CastilloNessuna valutazione finora
- Aparicio Et Al. 2015 Anfibios Del Valle de La Paz BoliviaDocumento19 pagineAparicio Et Al. 2015 Anfibios Del Valle de La Paz Boliviakitita2707Nessuna valutazione finora
- 163 185 RIA 38 Iberesia Castillana Frade y Bacelar 1931 Araneae Nemesiidae 11Documento23 pagine163 185 RIA 38 Iberesia Castillana Frade y Bacelar 1931 Araneae Nemesiidae 11Ezequiel Prieto LilloNessuna valutazione finora
- Terenotriccus ErythrurusDocumento4 pagineTerenotriccus ErythrurusMichel AlvaNessuna valutazione finora
- Musniv v1Documento18 pagineMusniv v1darioNessuna valutazione finora
- Laboratorios 8 y 9 Ento Parte LitzyDocumento9 pagineLaboratorios 8 y 9 Ento Parte LitzyDenis PérezNessuna valutazione finora
- Centrolenecondor 2012 Politecnica 303Documento13 pagineCentrolenecondor 2012 Politecnica 303lasotru743Nessuna valutazione finora
- Orca - Orcinus OrcaDocumento12 pagineOrca - Orcinus OrcaLanoca DeFresaNessuna valutazione finora
- VenadoDocumento28 pagineVenadoJuan Carlos Roa RoaNessuna valutazione finora
- Jabalí PDFDocumento26 pagineJabalí PDFanzaeduardo1Nessuna valutazione finora
- Tityus Cerroazul Lourenço (ScorpionesDocumento15 pagineTityus Cerroazul Lourenço (ScorpionesCarlos MoránNessuna valutazione finora
- Acaros Asociados A Vertebrados Montiel-Parra Et Al 2009Documento10 pagineAcaros Asociados A Vertebrados Montiel-Parra Et Al 2009Jesús Campos SerranoNessuna valutazione finora
- Nueva Especie Exotica de Mamifero en LaDocumento8 pagineNueva Especie Exotica de Mamifero en LaDiego MoralesNessuna valutazione finora
- Mamiferos de El SalvadorDocumento299 pagineMamiferos de El SalvadorCarlos HormazábalNessuna valutazione finora
- 05 NA-58 SalvadorDocumento6 pagine05 NA-58 SalvadorSergio Eduardo VillagraNessuna valutazione finora
- Grupo 16 Ex Aula 2Documento19 pagineGrupo 16 Ex Aula 2Jeremy RuizNessuna valutazione finora
- Biología y Morfología de Morpho Menelaus GodartiiDocumento9 pagineBiología y Morfología de Morpho Menelaus GodartiiRx SangaNessuna valutazione finora
- Anfibios TetraploidiaDocumento15 pagineAnfibios TetraploidiaIsabel CuadrosNessuna valutazione finora
- Claves NeuropteraDocumento10 pagineClaves NeuropteraFernando Ancco ValdiviaNessuna valutazione finora
- Caso Histórico Pez LeonDocumento12 pagineCaso Histórico Pez LeonMaria Teresa Carreon ZapiainNessuna valutazione finora
- Clave ColeopterosDocumento17 pagineClave ColeopterosDianita FoxNessuna valutazione finora
- ReptilesCuyabenoFinal PDFDocumento292 pagineReptilesCuyabenoFinal PDFivan lopezNessuna valutazione finora
- Monografia - Isa IsasiDocumento12 pagineMonografia - Isa IsasiIván Rolando Díaz SandovalNessuna valutazione finora
- Drosophila EcuadorDocumento13 pagineDrosophila EcuadorJOSE FRANCISCO FRANCO NAVIANessuna valutazione finora
- Anfibios CorreccionDocumento5 pagineAnfibios CorreccionLuis Fernando SáenzNessuna valutazione finora
- Loro RealDocumento24 pagineLoro Realrosm531Nessuna valutazione finora
- Clave Coleopteros PDFDocumento17 pagineClave Coleopteros PDFikkizodiacNessuna valutazione finora
- Ficha ACH Riama 66 72Documento7 pagineFicha ACH Riama 66 72Ana Maria MorenoNessuna valutazione finora
- NeuropteraDocumento22 pagineNeuropteraAndres ChuraNessuna valutazione finora
- The Sea Cucumber Echinopsolus Murrayi Théel, 1886 N. Comb. (Echinodermata: Holothuroidea) From The Southwestern Atlantic, Redescription of The Holotype and Description of New SpecimensDocumento6 pagineThe Sea Cucumber Echinopsolus Murrayi Théel, 1886 N. Comb. (Echinodermata: Holothuroidea) From The Southwestern Atlantic, Redescription of The Holotype and Description of New Specimensmonialicardozo63Nessuna valutazione finora
- 006a Bibliografía Invertebrados ParaguayDocumento188 pagine006a Bibliografía Invertebrados ParaguayJohn A. KochalkaNessuna valutazione finora
- 2012 PACE Perrito LlaneroDocumento36 pagine2012 PACE Perrito LlaneroLuca RoNessuna valutazione finora
- Parnass I UsDocumento4 pagineParnass I UsБренда ЛесамаNessuna valutazione finora
- Diferenciación de Ecotipos en El Perro de Agua EspañolDocumento5 pagineDiferenciación de Ecotipos en El Perro de Agua EspañolAna LopezNessuna valutazione finora
- Guia Reptiles CalakmulDocumento57 pagineGuia Reptiles CalakmulRoberto Rivera GuzmanNessuna valutazione finora
- Escolopendras en VenezuelaDocumento7 pagineEscolopendras en VenezuelaCarlos Ortíz SegoviaNessuna valutazione finora
- CATALOGO INIA - Gallinas PDFDocumento21 pagineCATALOGO INIA - Gallinas PDFElisabeth Rivas AlcaideNessuna valutazione finora
- Guia PecesDocumento11 pagineGuia PecesLaura Avila BarreraNessuna valutazione finora
- Razas Abejas FinalDocumento29 pagineRazas Abejas FinalMarcelo Andres Mendez Calbuyahue100% (1)
- Pica PDFDocumento10 paginePica PDFjavi17Nessuna valutazione finora
- Urraca - Pica Pica Linnaeus, 1758Documento19 pagineUrraca - Pica Pica Linnaeus, 1758pulpopulpoNessuna valutazione finora
- 010albaetal 2001spiraDocumento13 pagine010albaetal 2001spirapulpopulpoNessuna valutazione finora
- Pica Pica tcm30-200851Documento1 paginaPica Pica tcm30-200851pulpopulpoNessuna valutazione finora
- Urraca - Pica Pica Linnaeus, 1758Documento19 pagineUrraca - Pica Pica Linnaeus, 1758pulpopulpoNessuna valutazione finora
- Urraca tcm30-100122Documento2 pagineUrraca tcm30-100122pulpopulpoNessuna valutazione finora
- Milano Real tcm30-100076Documento2 pagineMilano Real tcm30-100076pulpopulpoNessuna valutazione finora
- PDF 79Documento28 paginePDF 79pulpopulpoNessuna valutazione finora
- Gyps Fulvus2Documento2 pagineGyps Fulvus2pulpopulpoNessuna valutazione finora
- TFGjuny ARCO SILVA - MACARENA MNDocumento37 pagineTFGjuny ARCO SILVA - MACARENA MNpulpopulpoNessuna valutazione finora
- Gyps Fulvus tcm30-200788Documento1 paginaGyps Fulvus tcm30-200788pulpopulpoNessuna valutazione finora
- Colaboracion GpsDocumento29 pagineColaboracion GpspulpopulpoNessuna valutazione finora
- MARM Ficha Técnica Del Buitre LeonadoDocumento2 pagineMARM Ficha Técnica Del Buitre LeonadotourismaldeadavilaNessuna valutazione finora
- 2.37. Especie A078. Gyps Fulvus (Buitre Leonado) : 2.37.1. Área de DistribuciónDocumento7 pagine2.37. Especie A078. Gyps Fulvus (Buitre Leonado) : 2.37.1. Área de DistribuciónpulpopulpoNessuna valutazione finora
- El Buitre Leonado Gyps Fulvus en Islas Del Mediterráneo: Colonización y Asentamiento en Las Islas BalearesDocumento10 pagineEl Buitre Leonado Gyps Fulvus en Islas Del Mediterráneo: Colonización y Asentamiento en Las Islas BalearespulpopulpoNessuna valutazione finora
- Gypful v4Documento31 pagineGypful v4pulpopulpoNessuna valutazione finora
- Halcón Peregrino - Falco Peregrinus Tunstall, 1771Documento29 pagineHalcón Peregrino - Falco Peregrinus Tunstall, 1771SERGIONessuna valutazione finora
- Halcon Peregrino (Falco)Documento2 pagineHalcon Peregrino (Falco)pulpopulpoNessuna valutazione finora
- Nocardia AsteroidesDocumento11 pagineNocardia AsteroidespulpopulpoNessuna valutazione finora
- Buitre Leonado tcm30-195030Documento3 pagineBuitre Leonado tcm30-195030pulpopulpoNessuna valutazione finora
- Fecundacion Erizos de MarDocumento6 pagineFecundacion Erizos de MarTomas GianniNessuna valutazione finora
- Orden AraneaeDocumento13 pagineOrden AraneaepulpopulpoNessuna valutazione finora
- Erizo de MarDocumento3 pagineErizo de MarpulpopulpoNessuna valutazione finora
- Centrostephanus Longispinus tcm7-272540 PDFDocumento58 pagineCentrostephanus Longispinus tcm7-272540 PDFpulpopulpoNessuna valutazione finora
- 9 Chlorophyta TextoDocumento5 pagine9 Chlorophyta TextoJuan Manuel Hernández HernándezNessuna valutazione finora
- E. ColiDocumento12 pagineE. ColiMaría RicoNessuna valutazione finora
- CastañoDocumento4 pagineCastañopulpopulpoNessuna valutazione finora
- Expansion Del Buitre Leonado Gyps Fulvus: Tamaño de La Poblacion y Parametros Reproductores en Un Area de Reciente ColonizacionDocumento8 pagineExpansion Del Buitre Leonado Gyps Fulvus: Tamaño de La Poblacion y Parametros Reproductores en Un Area de Reciente ColonizacionpulpopulpoNessuna valutazione finora
- Articulo Cientifico PDFDocumento6 pagineArticulo Cientifico PDFNeider Jose Campo HernandezNessuna valutazione finora
- Pensum AgroalimentariaDocumento16 paginePensum AgroalimentariaArgenis PoleoNessuna valutazione finora
- Alexander PreguntasDocumento4 pagineAlexander PreguntasALEXANDER JHONATAN MARIN FLORESNessuna valutazione finora
- Ndie U2 A2 BelgDocumento15 pagineNdie U2 A2 BelgEllen Lucge100% (1)
- ElaboraciondetortillasregionalesdesanmartinDocumento34 pagineElaboraciondetortillasregionalesdesanmartintoacaminhoNessuna valutazione finora
- Qué Es El AmoniacoDocumento2 pagineQué Es El AmoniacoGeovanny MaquilonNessuna valutazione finora
- Vitamina A B C D KDocumento3 pagineVitamina A B C D KCharly CosquillasNessuna valutazione finora
- NL20150925 PDFDocumento96 pagineNL20150925 PDFconstruccionestytNessuna valutazione finora
- Ejemplo de Dieta Hiperproteica para Vegetarianos Que Quieren Ganar MúsculoDocumento10 pagineEjemplo de Dieta Hiperproteica para Vegetarianos Que Quieren Ganar MúsculoAzucaNessuna valutazione finora
- Mi Proyecto de Melon CaseroDocumento12 pagineMi Proyecto de Melon CaseroDavid CastroNessuna valutazione finora
- Codigo de Colores y SeñalesDocumento35 pagineCodigo de Colores y SeñalesJohn CamposNessuna valutazione finora
- Polleria Campos Final AMALIADocumento46 paginePolleria Campos Final AMALIALeydy Kaori Sucasaca RamosNessuna valutazione finora
- Veterinary Focus - 2017 - 27.2.es PDFDocumento52 pagineVeterinary Focus - 2017 - 27.2.es PDFNicole SaucedoNessuna valutazione finora
- Plan de Contingencia EdasDocumento17 paginePlan de Contingencia EdasEvelyn Mallqui DiazNessuna valutazione finora
- El Agua en Los Alimentos (H2O)Documento4 pagineEl Agua en Los Alimentos (H2O)Nancy MamaniNessuna valutazione finora
- El Biohuerto Casero Un Pedacito Ecologico en Tu HogarDocumento9 pagineEl Biohuerto Casero Un Pedacito Ecologico en Tu HogarRosmery Celinda RiverosNessuna valutazione finora
- BacteriologiaDocumento30 pagineBacteriologiaAlvaro Israel Zapata VargasNessuna valutazione finora
- Anuario Cenem 2015 - 1Documento132 pagineAnuario Cenem 2015 - 1Diego SalazarNessuna valutazione finora
- C.N. - 6 - 2Documento5 pagineC.N. - 6 - 2Jose Angel Agualimpia MosqueraNessuna valutazione finora
- Investigación de La Acelga (Beta Vulgaris) Y Propuesta Gastronómica en El Cantón de Colta de La Provincia de ChimborazoDocumento166 pagineInvestigación de La Acelga (Beta Vulgaris) Y Propuesta Gastronómica en El Cantón de Colta de La Provincia de Chimborazojennifer roseroNessuna valutazione finora
- Tratamientos y Depuración de Las Aguas Residuales - Metcalf Eddy - Cap 7Documento49 pagineTratamientos y Depuración de Las Aguas Residuales - Metcalf Eddy - Cap 7mansocalula80% (5)
- Metabolismo Energetico UltimoDocumento30 pagineMetabolismo Energetico UltimoPorfirio Diaz Miron100% (1)
- Resolucion Administrativa #74-2012Documento37 pagineResolucion Administrativa #74-2012henry PilcoNessuna valutazione finora
- El Celo y Sus EtapasDocumento30 pagineEl Celo y Sus EtapasDamian GiovinazzoNessuna valutazione finora
- Analisis Foda de MexicoDocumento7 pagineAnalisis Foda de MexicoAnonymous yMn6VLwOE100% (3)
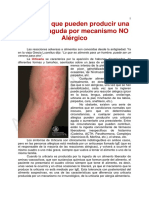
- Alimentos y Urticaria NO AlergicaDocumento3 pagineAlimentos y Urticaria NO AlergicaSelene MorningGloryNessuna valutazione finora
- El Laboratorio MicrobiologicoDocumento192 pagineEl Laboratorio MicrobiologicomarcelaNessuna valutazione finora
- Clasificación de Manejo de Residuos SolidosDocumento1 paginaClasificación de Manejo de Residuos SolidosKaro Alvarez Sanchez100% (2)
- Proyecto El TrapicheDocumento8 pagineProyecto El TrapicheLeopoldo Mendoza100% (1)
- Dieta BlandaDocumento7 pagineDieta BlandaVeronika RodriguezNessuna valutazione finora
- Periódico Del Plato Del Buen ComerDocumento9 paginePeriódico Del Plato Del Buen ComerAdan MendozaNessuna valutazione finora