Potrebbero piacerti anche
- I.J. Singh, Professor Department of Fishery Biology College of Fisheries, G.B.P.U.A.&T., PantnagarDocumento39 pagineI.J. Singh, Professor Department of Fishery Biology College of Fisheries, G.B.P.U.A.&T., Pantnagarjoshigauta100% (1)
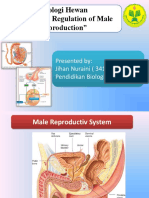
- Male ReproductiveDocumento10 pagineMale ReproductivevivekNessuna valutazione finora
- Breeding and Spawning of Fishes Role of Endocrine Gland PDFDocumento7 pagineBreeding and Spawning of Fishes Role of Endocrine Gland PDFSachin KhairnarNessuna valutazione finora
- Endocrine Physiology) 21. Female Reproductive Cycle - Ovulation - KeyDocumento1 paginaEndocrine Physiology) 21. Female Reproductive Cycle - Ovulation - Keyhasanatiya41Nessuna valutazione finora
- Gynecology Review: Abigail Elsie Dg. Castro, MD, Maed, Fpogs, Fpsuog August 11, 2017Documento198 pagineGynecology Review: Abigail Elsie Dg. Castro, MD, Maed, Fpogs, Fpsuog August 11, 2017Jojo Mendoza100% (1)
- Functional Anatomy of The Hypothalamicpituitarygonadal Axis andDocumento10 pagineFunctional Anatomy of The Hypothalamicpituitarygonadal Axis andEvaNessuna valutazione finora
- Anfis Sistem Reproduksi Pria Dan WanitaDocumento62 pagineAnfis Sistem Reproduksi Pria Dan WanitaLili HamdianaNessuna valutazione finora
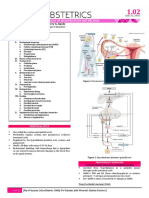
- OB 1 - 1.02 Physiology of Menstruation and Decidua PDFDocumento11 pagineOB 1 - 1.02 Physiology of Menstruation and Decidua PDFMDreamerNessuna valutazione finora
- Female Endocrinology - AnimalDocumento37 pagineFemale Endocrinology - AnimalLevi Junho SongNessuna valutazione finora
- Reproductive Physiology PDFDocumento70 pagineReproductive Physiology PDFDrbee10Nessuna valutazione finora
- Endocrine System Regulates Animal PhysiologyDocumento9 pagineEndocrine System Regulates Animal PhysiologyFlor SagnipNessuna valutazione finora
- Endocrinology:: By: DR Suyash SharmaDocumento186 pagineEndocrinology:: By: DR Suyash SharmaaalipuriaNessuna valutazione finora
- 6-TheR 608# Hormones & Reprod-IIDocumento15 pagine6-TheR 608# Hormones & Reprod-IIHamza HassanNessuna valutazione finora
- First Aid For The OBS&GYN Clerkship CH 14 (Menstruation)Documento4 pagineFirst Aid For The OBS&GYN Clerkship CH 14 (Menstruation)黃芳昌Nessuna valutazione finora
- PUBERTY: THE STAGES OF HUMAN DEVELOPMENTDocumento62 paginePUBERTY: THE STAGES OF HUMAN DEVELOPMENTvaluevaNessuna valutazione finora
- Hypothalamus Pituitary and Gonad AxisDocumento6 pagineHypothalamus Pituitary and Gonad Axisalok nayakNessuna valutazione finora
- Regulacion de Epo Journal Fisiol 2012Documento8 pagineRegulacion de Epo Journal Fisiol 2012Hugo HectorNessuna valutazione finora
- Kuliah 2.1 - SIKLUS MENSTRUASIDocumento12 pagineKuliah 2.1 - SIKLUS MENSTRUASIErviana Dwi NurhidayatiNessuna valutazione finora
- The Basics of Promoting Fish Maturation and SpawningDocumento11 pagineThe Basics of Promoting Fish Maturation and Spawningkatral1968Nessuna valutazione finora
- Endocrine System - Part 1 (Robbins)Documento28 pagineEndocrine System - Part 1 (Robbins)sarguss14100% (2)
- L26 PDFDocumento44 pagineL26 PDFMiles HuiNessuna valutazione finora
- Female Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeDocumento56 pagineFemale Reproductive Physiology: Pradip Kumar Das, Joydip Mukherjee, and Dipak BanerjeeVFJulianNessuna valutazione finora
- (TLOY) OB - Endometrial and Ovarian CycleDocumento6 pagine(TLOY) OB - Endometrial and Ovarian CycleJAMICA ROMAINE AMBIONNessuna valutazione finora
- Endocrine Day 1 TemplateDocumento17 pagineEndocrine Day 1 TemplateMikeNessuna valutazione finora
- Review of Endocrine SystemDocumento32 pagineReview of Endocrine SystemJames FNessuna valutazione finora
- Ovarian Life Cycle: Jimmy Yanuar AnnasDocumento55 pagineOvarian Life Cycle: Jimmy Yanuar AnnasFellita Ratri ANessuna valutazione finora
- HormonesDocumento29 pagineHormonesDivya Thuruthiyil RamachandranNessuna valutazione finora
- Bio 102 - Chapter 18: Endocrine Organs: Neurohypophysis Median Eminence Infundibular Stalk Pars NervosaDocumento7 pagineBio 102 - Chapter 18: Endocrine Organs: Neurohypophysis Median Eminence Infundibular Stalk Pars NervosaAlly YeeNessuna valutazione finora
- Endocrine SystemDocumento7 pagineEndocrine SystemIvy Jana BelenNessuna valutazione finora
- Topic3.2 Physiology Hypothalamus and Pituitary HormonesDocumento4 pagineTopic3.2 Physiology Hypothalamus and Pituitary HormonesRen AlvNessuna valutazione finora
- Human Reproductive Endocrinology: Biol/Bmsc 114Documento27 pagineHuman Reproductive Endocrinology: Biol/Bmsc 114Marcus EbertNessuna valutazione finora
- ImmunologyDocumento2 pagineImmunologyDebapriya HazraNessuna valutazione finora
- Endokrinologi Reproduksi: Bayu Kurniawan, DRDocumento48 pagineEndokrinologi Reproduksi: Bayu Kurniawan, DRdocbay100% (1)
- SBM Reproduction NotesDocumento73 pagineSBM Reproduction NotesAhra100% (2)
- ENDORCRINE SYSTEM - Review Anaphy NotesDocumento10 pagineENDORCRINE SYSTEM - Review Anaphy NotesJae TyNessuna valutazione finora
- Brown & McNeillyDocumento9 pagineBrown & McNeillyFérielNessuna valutazione finora
- Lab Report Assistant Endocrine SystemDocumento7 pagineLab Report Assistant Endocrine SystemJohn Louis AguilaNessuna valutazione finora
- Male Reproductive HormonesDocumento29 pagineMale Reproductive HormonesAjit ShahNessuna valutazione finora
- Reproductive Physiology: RP Lecture II Series The FemaleDocumento27 pagineReproductive Physiology: RP Lecture II Series The FemaleAsad JiwaniNessuna valutazione finora
- Desarrollo Puberal, Pediatr Rev. 2016 Jul 37 (7) 292-300Documento11 pagineDesarrollo Puberal, Pediatr Rev. 2016 Jul 37 (7) 292-300RUBY JOJOANessuna valutazione finora
- Human Preproduction Master CopyDocumento72 pagineHuman Preproduction Master CopyShalini JhaNessuna valutazione finora
- OB Ozamiz GroupDocumento26 pagineOB Ozamiz GroupJb RosillosaNessuna valutazione finora
- 1st Lecture - Hypothalamus Pituitary Gonadal Axis - Reproductive PhysiologyDocumento8 pagine1st Lecture - Hypothalamus Pituitary Gonadal Axis - Reproductive PhysiologyBHUWAN BASKOTANessuna valutazione finora
- Gland Stimulation of Hormones Produced Action Gland: Fsh/Lh-Gonads ActhDocumento2 pagineGland Stimulation of Hormones Produced Action Gland: Fsh/Lh-Gonads ActhshirleyNessuna valutazione finora
- Platelet Development: An Overview of MegakaryopoiesisDocumento5 paginePlatelet Development: An Overview of MegakaryopoiesisPaul LesterNessuna valutazione finora
- Male Reproduction Hormone RegulationDocumento8 pagineMale Reproduction Hormone RegulationocktiNessuna valutazione finora
- Endocrine System Animal ScienceDocumento15 pagineEndocrine System Animal ScienceJairus LampanoNessuna valutazione finora
- P.10 HYPOTHALAMUS AND PITUITARY GLAND PART 1 and 2 (DR. ORIBIO) 02-05-18Documento7 pagineP.10 HYPOTHALAMUS AND PITUITARY GLAND PART 1 and 2 (DR. ORIBIO) 02-05-18Raymund Dan AldabaNessuna valutazione finora
- Hormone Action in The Mammary Gland - C Brisken and B O Malley 2010Documento15 pagineHormone Action in The Mammary Gland - C Brisken and B O Malley 2010tostonasNessuna valutazione finora
- Javern M. Supnet RNDocumento16 pagineJavern M. Supnet RNGenghis SupnetNessuna valutazione finora
- HY EndocrineoeDocumento62 pagineHY EndocrineoebrownsmilansNessuna valutazione finora
- HY EndocrineDocumento61 pagineHY EndocrineKiranNessuna valutazione finora
- Endocrine System of FishesDocumento5 pagineEndocrine System of FishesmonjitpaulNessuna valutazione finora
- Phys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESDocumento7 paginePhys 6.1 and 6.2 Ant Pituitary GH and Post Pituitary NOTESsine summsun PradhanNessuna valutazione finora
- Male Central Hypogonadism in Paediatrics - The Relevance of Follicle-Stimulating Hormone and Sertoli Cell MarkersDocumento5 pagineMale Central Hypogonadism in Paediatrics - The Relevance of Follicle-Stimulating Hormone and Sertoli Cell MarkersScott LoveNessuna valutazione finora
- Endocrine Glands - 1st - ChapterDocumento12 pagineEndocrine Glands - 1st - Chaptervarun kumarNessuna valutazione finora
- Anfis Endokrin S2 (Compatibility Mode) .CompressedDocumento66 pagineAnfis Endokrin S2 (Compatibility Mode) .CompressedAmiliya EmilNessuna valutazione finora
- 1892 Fdoc PDFDocumento15 pagine1892 Fdoc PDFLavender B MatjeleNessuna valutazione finora
- Care of The Patient With Complex Endocrine Disorders Endocrine SystemDocumento11 pagineCare of The Patient With Complex Endocrine Disorders Endocrine SystemgoodgoNessuna valutazione finora
- Introduction to Clinical Reproductive EndocrinologyDa EverandIntroduction to Clinical Reproductive EndocrinologyValutazione: 1 su 5 stelle1/5 (1)
- Lumify Tablet Compatibility: Screen Size Weight Approximate Continuous Scan Time Connectivity Options Internal StorageDocumento1 paginaLumify Tablet Compatibility: Screen Size Weight Approximate Continuous Scan Time Connectivity Options Internal StorageUmi RehanNessuna valutazione finora
- Full Paper: DEBRIS SEL Aeromonas Hydrophila PADA LELE DUMBO (Clarias SP.)Documento8 pagineFull Paper: DEBRIS SEL Aeromonas Hydrophila PADA LELE DUMBO (Clarias SP.)Umi RehanNessuna valutazione finora
- Soal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Documento7 pagineSoal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Umi RehanNessuna valutazione finora
- Aquagenome Review DraftDocumento26 pagineAquagenome Review DraftUmi RehanNessuna valutazione finora
- Soal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Documento7 pagineSoal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Umi RehanNessuna valutazione finora
- Using Copepod Oithona sp as Live Feed for Grouper LarvaeDocumento8 pagineUsing Copepod Oithona sp as Live Feed for Grouper LarvaeMuh. AriefNessuna valutazione finora
- Soal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Documento7 pagineSoal Osn Biologi Tingkat Kabupaten Kota Tahun 2015Umi RehanNessuna valutazione finora
- Jones Pub 33Documento6 pagineJones Pub 33Umi RehanNessuna valutazione finora
- Yuhuuuuuuuuuuuuuuuuuuuuuuu Kepada Yth. Ketua LPPMDocumento1 paginaYuhuuuuuuuuuuuuuuuuuuuuuuu Kepada Yth. Ketua LPPMUmi RehanNessuna valutazione finora
- Kel 8. EAA in Shrimp Culture at ThailandDocumento53 pagineKel 8. EAA in Shrimp Culture at ThailandUmi RehanNessuna valutazione finora
- Pathophysiology of Pulmonary Embolism (Loria.J)Documento2 paginePathophysiology of Pulmonary Embolism (Loria.J)Justine Mae Loria0% (1)
- False 2. True 3. True 4. True 5. False 6. False 7. False 8. True 9. True 10. TrueDocumento43 pagineFalse 2. True 3. True 4. True 5. False 6. False 7. False 8. True 9. True 10. TrueKero KeropiNessuna valutazione finora
- Cuttings Propagation of Labisia Pumila (Kacip Fatimah)Documento7 pagineCuttings Propagation of Labisia Pumila (Kacip Fatimah)Felix VioletaNessuna valutazione finora
- Anatomy and Physiology of LarynxDocumento37 pagineAnatomy and Physiology of Larynxsuci100% (1)
- Cardiovascular and Pulmonary Physical Therapy, 2nd EditionDocumento439 pagineCardiovascular and Pulmonary Physical Therapy, 2nd EditionJason100% (3)
- NUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityDocumento3 pagineNUR11O1 Integrated Human Anatomy and Physiology Department of Biology Institute of Arts and Sciences Far Eastern UniversityPrancheska Abigayle Peneyra SantiagoNessuna valutazione finora
- Cells 10 02857 v2Documento33 pagineCells 10 02857 v2DiegoNessuna valutazione finora
- Shs Stem - Biology 1: Quarter 1 - Module 14Documento21 pagineShs Stem - Biology 1: Quarter 1 - Module 14Grace RellosoNessuna valutazione finora
- 3 Aquatic PlantsDocumento43 pagine3 Aquatic PlantsMhyna klauNessuna valutazione finora
- Virgen Milagrosa University Foundation College of Radiologic Technology Human Anatomy and PhysiologyDocumento2 pagineVirgen Milagrosa University Foundation College of Radiologic Technology Human Anatomy and PhysiologyJas-Stephanie MarcoNessuna valutazione finora
- Hormone Practice Test: StudentDocumento17 pagineHormone Practice Test: StudentKayne SuratosNessuna valutazione finora
- Physiological:biological Psychology NotesDocumento36 paginePhysiological:biological Psychology NotesjeynNessuna valutazione finora
- Rat Dissection Guide: Including Pregnant FemaleDocumento16 pagineRat Dissection Guide: Including Pregnant FemaleMAVIE GOMEZNessuna valutazione finora
- LC Bright BrochureDocumento1 paginaLC Bright Brochureapi-528907705Nessuna valutazione finora
- Stages of Development BrainDocumento6 pagineStages of Development BrainMuhammad ZubairNessuna valutazione finora
- Physio Ex 3-01Documento8 paginePhysio Ex 3-01edie tyler100% (4)
- (Unit) Master Ekatalog CMC 2022Documento75 pagine(Unit) Master Ekatalog CMC 2022Promkes PuskSusut2Nessuna valutazione finora
- Feature Article The Neurobiology of Thought: The Groundbreaking Discoveries of Patricia Goldman-Rakic 1937 - 2003Documento13 pagineFeature Article The Neurobiology of Thought: The Groundbreaking Discoveries of Patricia Goldman-Rakic 1937 - 2003Adelina MarițiuNessuna valutazione finora
- Ciliates & SporozoaDocumento50 pagineCiliates & SporozoaBalach JanNessuna valutazione finora
- 15 - Digestive System Anatomy and Physiology SeriesDocumento101 pagine15 - Digestive System Anatomy and Physiology SeriesIsaac KipkemoiNessuna valutazione finora
- T H T Y L C: HE AND AI IN UNG HannelDocumento10 pagineT H T Y L C: HE AND AI IN UNG Hannelveerasekhar.chNessuna valutazione finora
- 11B2 Biological ClassificationDocumento52 pagine11B2 Biological Classificationaaravshah1256Nessuna valutazione finora
- Krok 1 - 2018 (General Medicine) - EneutronDocumento57 pagineKrok 1 - 2018 (General Medicine) - EneutronHarsh NimavatNessuna valutazione finora
- Diadynamic CurrentDocumento44 pagineDiadynamic Currentbikedet268Nessuna valutazione finora
- Hematopoietic Agents and Hematinics: PharmacologyDocumento9 pagineHematopoietic Agents and Hematinics: PharmacologyAbi SulitNessuna valutazione finora
- Cytokines, Chemokines and Their Receptors - Madame Curie Bioscience Database - NCBI BookshelfDocumento25 pagineCytokines, Chemokines and Their Receptors - Madame Curie Bioscience Database - NCBI BookshelfRezki IsharNessuna valutazione finora
- Circadian Rhythms and Biological Clocks Part B PDFDocumento389 pagineCircadian Rhythms and Biological Clocks Part B PDFLeoNessuna valutazione finora
- Enzymes ReviewerDocumento2 pagineEnzymes ReviewerAshley MalolotNessuna valutazione finora
- Biology 1: Learning Activity SheetDocumento184 pagineBiology 1: Learning Activity SheetJohn joseph Ursulum100% (1)
- Disp Sets and Filler OverviewDocumento1 paginaDisp Sets and Filler OverviewAnil Kumar kakaraparthiNessuna valutazione finora