Potrebbero piacerti anche
- UBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatDocumento18 pagineUBA - CBC - Biologia - Resumen Libritos 2, 3, 4 y 6 - CatJuanJoseNessuna valutazione finora
- RMN - 2D Varios Diapos PDFDocumento32 pagineRMN - 2D Varios Diapos PDFquimica4dfNessuna valutazione finora
- Organización y Funciones de La Célula Eucariota OkDocumento5 pagineOrganización y Funciones de La Célula Eucariota Oknaziantonio1Nessuna valutazione finora
- Organelos y Sus Funciones - LHG - Subir Como Material 28 Sep 2021Documento57 pagineOrganelos y Sus Funciones - LHG - Subir Como Material 28 Sep 2021AGUILAR CALDERÓN OSVALDONessuna valutazione finora
- Anato 1Documento61 pagineAnato 1NALLELYNessuna valutazione finora
- 1-I. Procesos BiologicosDocumento10 pagine1-I. Procesos BiologicosLuis Ed Navarro IriarteNessuna valutazione finora
- U2 Act 2 Estructura Fisica de La CelulaDocumento15 pagineU2 Act 2 Estructura Fisica de La CelulaKAROL GUTIERREZ MADRIGALNessuna valutazione finora
- Organelos CelularesDocumento9 pagineOrganelos CelularesVictor Gonzalez Garcia0% (1)
- Biología Clase 27.04Documento18 pagineBiología Clase 27.04Camii MariqueoNessuna valutazione finora
- Célula AnimalDocumento17 pagineCélula AnimalJUDITH ALONDRA TORRES HERNANDEZNessuna valutazione finora
- Estructurasyorganelosdelacelula 10 Bio PDFDocumento7 pagineEstructurasyorganelosdelacelula 10 Bio PDFPablo HernàndezNessuna valutazione finora
- Estructura y Fisiología CelularDocumento23 pagineEstructura y Fisiología CelularValentina PeraltaNessuna valutazione finora
- La CélulaDocumento25 pagineLa CélulaNîltøn ĆhNessuna valutazione finora
- Organelos Que Trasportan y Crean ProteinasDocumento7 pagineOrganelos Que Trasportan y Crean ProteinasLetty Coronel100% (2)
- Laboratorio Fundamento Teórico Tema IX Cel AnimalDocumento12 pagineLaboratorio Fundamento Teórico Tema IX Cel AnimalGabriela Noemi Cayo ChoqueNessuna valutazione finora
- La CélulaDocumento14 pagineLa CélulaDeiv PerezNessuna valutazione finora
- Informe de LaboratorioDocumento4 pagineInforme de LaboratorioAlejandro AliendreNessuna valutazione finora
- G1 Estructura y Funcion CelularDocumento14 pagineG1 Estructura y Funcion CelularYaritza AhumadaNessuna valutazione finora
- Copia de Tema 6 ORGANIZACION CELULARDocumento53 pagineCopia de Tema 6 ORGANIZACION CELULARAMIRA MOHAMED EL HARRAKNessuna valutazione finora
- Manual Anatomía 2022-2023Documento318 pagineManual Anatomía 2022-2023sebastianerdos78Nessuna valutazione finora
- Células ProcariotasDocumento3 pagineCélulas ProcariotasAnonymous gmUWuaNessuna valutazione finora
- Celula TareaDocumento3 pagineCelula TareaMauricio EduardoNessuna valutazione finora
- Bibliografia Cel Procarioticas y Eucarioticas Clase 2Documento8 pagineBibliografia Cel Procarioticas y Eucarioticas Clase 2Nancy BertucciNessuna valutazione finora
- Organización CelularDocumento42 pagineOrganización CelularArturo Diaz PerezNessuna valutazione finora
- Citoplasma CelularDocumento19 pagineCitoplasma CelularAlisson Caicedo100% (2)
- Conferencia Citoplasma Completo OkDocumento60 pagineConferencia Citoplasma Completo OkPawan Alvarez PerezNessuna valutazione finora
- Guia Observación Celular BMS1100Documento10 pagineGuia Observación Celular BMS1100DIBL 96Nessuna valutazione finora
- Tema 3 Estructura y Función CelularDocumento8 pagineTema 3 Estructura y Función CelularJorge GarcíaNessuna valutazione finora
- Clase Tipos de CelulasDocumento35 pagineClase Tipos de CelulasAld Jann Carlo Trigozo Alava100% (1)
- Clase 5 - Procariota y EucariotaDocumento34 pagineClase 5 - Procariota y EucariotaPablo Latoja GómezNessuna valutazione finora
- Características de La Célula Animal y VegetalDocumento13 pagineCaracterísticas de La Célula Animal y VegetalJehison Figueroa100% (2)
- Tema 2. La Célula.Documento19 pagineTema 2. La Célula.Natalia Milla LopezNessuna valutazione finora
- CITOLOGIADocumento6 pagineCITOLOGIAestefany villcaNessuna valutazione finora
- Biologia Celular 4Documento12 pagineBiologia Celular 4Dolores de Jesús Marvin YahirNessuna valutazione finora
- 2023 Guía 1 Células y Estructuras CelularesDocumento12 pagine2023 Guía 1 Células y Estructuras CelularesclaudiaNessuna valutazione finora
- Taller Organulos CELULARESDocumento15 pagineTaller Organulos CELULARESFerney PalaciosNessuna valutazione finora
- La CélulaDocumento42 pagineLa CélulaEduardo Asael Tlaxalo RamirezNessuna valutazione finora
- Estructura Física de La CelulaDocumento10 pagineEstructura Física de La CelulaIvan Flores FloresNessuna valutazione finora
- Nivel CelularDocumento16 pagineNivel CelularDonnie Barzola de la ONessuna valutazione finora
- OrganelasDocumento4 pagineOrganelasEliana SolanoNessuna valutazione finora
- La Celula y Elementos ConstitutivosDocumento45 pagineLa Celula y Elementos ConstitutivosBeth DiazNessuna valutazione finora
- Citologia VegetalDocumento7 pagineCitologia VegetalkathehernandezNessuna valutazione finora
- Resumen 1 Cel y Tej - Met Est Del 1er Semestre MedicinaDocumento18 pagineResumen 1 Cel y Tej - Met Est Del 1er Semestre MedicinaDayan CespedesNessuna valutazione finora
- Taller Tren CelularDocumento3 pagineTaller Tren CelularEfren D. DiazNessuna valutazione finora
- Celulas Procariotas y EucariotasDocumento9 pagineCelulas Procariotas y EucariotasEstefany PeraltaNessuna valutazione finora
- CelulaDocumento7 pagineCelulagerardo mendozaNessuna valutazione finora
- Tarea 2 de BiologiaDocumento7 pagineTarea 2 de BiologiayerlinaNessuna valutazione finora
- Orgamelos (Biologia)Documento10 pagineOrgamelos (Biologia)Eduardo ManzanoNessuna valutazione finora
- Material Audiovisual CélulaDocumento25 pagineMaterial Audiovisual CélulaCristian MendozaNessuna valutazione finora
- CELULADocumento108 pagineCELULAalejandraNessuna valutazione finora
- Actividad de Repaso Procesos Biologicos OctavoDocumento7 pagineActividad de Repaso Procesos Biologicos OctavoSANDRANessuna valutazione finora
- Nivel CelularDocumento12 pagineNivel Celularyarotd sotoNessuna valutazione finora
- Microbiología Médica Cap 2Documento4 pagineMicrobiología Médica Cap 2Carlos Proaño Salmon100% (1)
- 03 Estructura y Función de Los OrganelosDocumento3 pagine03 Estructura y Función de Los OrganelosEsmeralda López Toro0% (1)
- La CelulaDocumento16 pagineLa CelulaStephany Mariela Espinoza SachaNessuna valutazione finora
- Wuolah Free App 1694792084005Documento95 pagineWuolah Free App 1694792084005Ángela PerezNessuna valutazione finora
- Biología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularDa EverandBiología Molecular y Celular: Una guía introductoria para aprender Biología Celular y MolecularNessuna valutazione finora
- Análisis funcional y microbiológico de derivados lácteos y cárnicosDa EverandAnálisis funcional y microbiológico de derivados lácteos y cárnicosNessuna valutazione finora
- Las células, Constructoras de vida: Cells: Constructing Living ThingsDa EverandLas células, Constructoras de vida: Cells: Constructing Living ThingsValutazione: 3.5 su 5 stelle3.5/5 (2)
- Arteria Aorta y Sus RamasDocumento4 pagineArteria Aorta y Sus RamasNicole Antonella RiosNessuna valutazione finora
- Irrigación de La Glándula TiroidesDocumento4 pagineIrrigación de La Glándula TiroidesNicole Antonella RiosNessuna valutazione finora
- Aparato RespiratorioDocumento22 pagineAparato RespiratorioNicole Antonella RiosNessuna valutazione finora
- Regiones Topograficas y Aponeurosis Del Miembro SuperiorDocumento8 pagineRegiones Topograficas y Aponeurosis Del Miembro SuperiorNicole Antonella RiosNessuna valutazione finora
- INTRODUCCIÓNDocumento10 pagineINTRODUCCIÓNNicole Antonella RiosNessuna valutazione finora
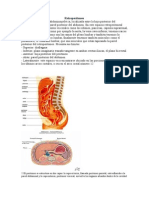
- RetroperitoneoDocumento2 pagineRetroperitoneoNicole Antonella RiosNessuna valutazione finora
- Hegemonía y Medios de ComunicaciónDocumento2 pagineHegemonía y Medios de ComunicaciónNicole Antonella RiosNessuna valutazione finora
- Anatomía Del CorazónDocumento9 pagineAnatomía Del CorazónNicole Antonella RiosNessuna valutazione finora
- Vascularizacion e Inervacion Del Miembro SuperiorDocumento27 pagineVascularizacion e Inervacion Del Miembro SuperiorFrancisco Navarrete Acuña100% (1)
- Regiones y Fosas ComunesDocumento3 pagineRegiones y Fosas Comunesluis22232Nessuna valutazione finora
- Actividades TP2 PDFDocumento3 pagineActividades TP2 PDFNicole Antonella RiosNessuna valutazione finora
- Resumen Anatomia ImagenologicaDocumento2 pagineResumen Anatomia ImagenologicaNicole Antonella RiosNessuna valutazione finora
- Triángulo FemoralDocumento2 pagineTriángulo FemoralNicole Antonella RiosNessuna valutazione finora
- Topografia Miembro SuperiorDocumento3 pagineTopografia Miembro SuperiorNicole Antonella Rios0% (1)
- AlcoholismoDocumento43 pagineAlcoholismoelchicharoteNessuna valutazione finora
- Músculos Del Miembro InferiorDocumento20 pagineMúsculos Del Miembro InferiorNicole Antonella RiosNessuna valutazione finora
- Topografia Miembro SuperiorDocumento3 pagineTopografia Miembro SuperiorNicole Antonella Rios0% (1)
- Librito 2,3,4 y 6Documento23 pagineLibrito 2,3,4 y 6Nicole Antonella RiosNessuna valutazione finora
- Resumen Biologia CelularDocumento43 pagineResumen Biologia CelularNicole Antonella RiosNessuna valutazione finora
- Librito 7,8,9y10Documento19 pagineLibrito 7,8,9y10Nicole Antonella RiosNessuna valutazione finora
- Librito 2,3,4 y 6Documento23 pagineLibrito 2,3,4 y 6Nicole Antonella RiosNessuna valutazione finora
- Librito 4 y 5Documento8 pagineLibrito 4 y 5Nicole Antonella RiosNessuna valutazione finora
- Librito 11,12 y 13Documento32 pagineLibrito 11,12 y 13Nicole Antonella RiosNessuna valutazione finora
- Librito 4 y 5Documento8 pagineLibrito 4 y 5Nicole Antonella RiosNessuna valutazione finora
- Librito 2,3,4 y 6Documento23 pagineLibrito 2,3,4 y 6Nicole Antonella RiosNessuna valutazione finora
- Biotecnologia de Hongos 1Documento16 pagineBiotecnologia de Hongos 1Katiuska MercadoNessuna valutazione finora
- Cultivo de CacahuateDocumento18 pagineCultivo de CacahuateNewsefiNessuna valutazione finora
- Aplicación de Aceites EsencialesDocumento10 pagineAplicación de Aceites EsencialesLulú DelatNessuna valutazione finora
- Manual de Bacteriologia y VirologiaDocumento88 pagineManual de Bacteriologia y VirologiaI.E.S.T.P. HIPOLITO UNANUE100% (1)
- Módulo 4 6° Básico FotosíntesisDocumento23 pagineMódulo 4 6° Básico Fotosíntesiscarla olivaNessuna valutazione finora
- Reproduccion de Los Seres VivosDocumento134 pagineReproduccion de Los Seres VivosChristian Raul Jorgensen100% (2)
- GUÍA DIDACTICA - 7 Ciencias NaturalesDocumento8 pagineGUÍA DIDACTICA - 7 Ciencias Naturaleslinaalexandra correaNessuna valutazione finora
- GUIA DE LABORATORIO No 5 - FITOQUIMICADocumento9 pagineGUIA DE LABORATORIO No 5 - FITOQUIMICAnicolas rodriguez valeroNessuna valutazione finora
- ADRIAN GALICIA SERRANO 5TO C DESARROLLO SUSTENTABLE Identificar Los Conceptos de Ecosistema, Los Elementos Que Lo Integran y Reconociendo El Papel Que Tienen Mediante Investigación BibliográficaDocumento9 pagineADRIAN GALICIA SERRANO 5TO C DESARROLLO SUSTENTABLE Identificar Los Conceptos de Ecosistema, Los Elementos Que Lo Integran y Reconociendo El Papel Que Tienen Mediante Investigación Bibliográficaw3stNessuna valutazione finora
- TE DE ESTIERCOl QuçomicaDocumento9 pagineTE DE ESTIERCOl QuçomicaDiego Rivas100% (1)
- Curso CONTR1Documento0 pagineCurso CONTR1Chihiro ChapulNessuna valutazione finora
- Evaluación de Competencia CurricularDocumento14 pagineEvaluación de Competencia CurricularElsa Fátima Ordi GlezNessuna valutazione finora
- CONICET Digital Nro. ADocumento58 pagineCONICET Digital Nro. AAdriana ChuliberNessuna valutazione finora
- Brochure Produccion Fresas HidroponicoDocumento20 pagineBrochure Produccion Fresas HidroponicoElnegro NegroNessuna valutazione finora
- Tema 1 Introduccion Al Estudio de La Anatomía y Morfología VegetalDocumento8 pagineTema 1 Introduccion Al Estudio de La Anatomía y Morfología VegetalJosé Justiniano Sánchez CárdenasNessuna valutazione finora
- Biologia-Partes de La Célula Animal y VegetalDocumento5 pagineBiologia-Partes de La Célula Animal y VegetalnelsondiezgarciaNessuna valutazione finora
- Enfermedades de Los CitricosDocumento4 pagineEnfermedades de Los CitricosGian Carlos Silva DiazNessuna valutazione finora
- Actividades de ObservacionDocumento2 pagineActividades de ObservacionNatalia LizarazoNessuna valutazione finora
- Actividad 4Documento5 pagineActividad 4Jennifer Vásquez67% (3)
- Apunte Tdai 2023Documento108 pagineApunte Tdai 2023Candela PereyraNessuna valutazione finora
- Manual de Densificación de La MaderaDocumento104 pagineManual de Densificación de La MaderaWilder R. Quispe RojasNessuna valutazione finora
- Actividades Mes MarzoDocumento26 pagineActividades Mes MarzoJorge FloresNessuna valutazione finora
- 260 Postulates Dynamics of Time by Jose Arguelles - En.esDocumento75 pagine260 Postulates Dynamics of Time by Jose Arguelles - En.esDiego LanconNessuna valutazione finora
- JP EcosistemasDocumento10 pagineJP Ecosistemaskeyvelin ruiz bonillaNessuna valutazione finora
- Realizar Una Investigación Documental de La Unidad 1. Introducción A La HolericulturaDocumento15 pagineRealizar Una Investigación Documental de La Unidad 1. Introducción A La Holericulturadaniel davidNessuna valutazione finora
- LechugaDocumento19 pagineLechugaCrysthelCardenasVillafuerteNessuna valutazione finora
- Guía Profe Sandra InsuastyDocumento2 pagineGuía Profe Sandra InsuastyVaneNessuna valutazione finora
- El Cosmos 2Documento47 pagineEl Cosmos 2Emmanuel AlexanderNessuna valutazione finora
- Cta4 Unidad 3Documento7 pagineCta4 Unidad 3Bruce Mirko Apaza BejarNessuna valutazione finora
- Belleza y Salud Tcm7-238480Documento8 pagineBelleza y Salud Tcm7-238480Faustino UrangaNessuna valutazione finora