Potrebbero piacerti anche
- DIY Home Remedy Recipes PDFDocumento6 pagineDIY Home Remedy Recipes PDFKarina Marcucci100% (1)
- The Witches 1996-1997 PDFDocumento50 pagineThe Witches 1996-1997 PDFNokatinNessuna valutazione finora
- Diet Weight GainDocumento2 pagineDiet Weight GainLalji Yadav100% (2)
- Food - Fruits, Vegetables & NutsDocumento58 pagineFood - Fruits, Vegetables & NutsThe 18th Century Material Culture Resource Center100% (3)
- Tree Plantation ProjectDocumento7 pagineTree Plantation ProjectNeglarized the world100% (2)
- Boiler ProblemsDocumento59 pagineBoiler ProblemsDharmendra BurraNessuna valutazione finora
- The Ultimate Aldi Grocery ListDocumento1 paginaThe Ultimate Aldi Grocery ListHallie AllenNessuna valutazione finora
- The Romantic Symbolism of TreesDocumento11 pagineThe Romantic Symbolism of TreesSol AbarcaNessuna valutazione finora
- Muscle and Weight Gain Diet PDFDocumento11 pagineMuscle and Weight Gain Diet PDFCavalera Max100% (1)
- Biotechnology 101 A Beginner's Guide To Modern Agricultural TechniquesDocumento43 pagineBiotechnology 101 A Beginner's Guide To Modern Agricultural TechniquesMaria Teresa V. DesturaNessuna valutazione finora
- Bioactive Endophytes Warrant Intensified Exploration and ConservationDocumento4 pagineBioactive Endophytes Warrant Intensified Exploration and ConservationNurul Robiatul AdawiyahNessuna valutazione finora
- Bacteriology RELACTTION - PLANT-BACTERIADocumento2 pagineBacteriology RELACTTION - PLANT-BACTERIAbiojacd5195Nessuna valutazione finora
- Differences Tight Loose Cultures 509Documento6 pagineDifferences Tight Loose Cultures 509salparNessuna valutazione finora
- Hox10 and Hox11 Genes Are Required To Globally Pattern The Mammalian SkeletonDocumento6 pagineHox10 and Hox11 Genes Are Required To Globally Pattern The Mammalian SkeletongomaNessuna valutazione finora
- Becker Ritt 2007Documento13 pagineBecker Ritt 2007Jucilene PereiraNessuna valutazione finora
- Zheng Et Al., 2013Documento13 pagineZheng Et Al., 2013jocyeoNessuna valutazione finora
- Science 1197754Documento5 pagineScience 1197754Josh TingNessuna valutazione finora
- 426 FullDocumento13 pagine426 FullJuan Carlos RedrujoNessuna valutazione finora
- Che Regeneration Rap2 6 jrg21 2006Documento18 pagineChe Regeneration Rap2 6 jrg21 2006Mariana GalgenNessuna valutazione finora
- CBO9780511902451A015Documento20 pagineCBO9780511902451A015Veronica B MarinaNessuna valutazione finora
- Short Communication Screening of Quorum Quenching Activity of Bacteria Isolated From Ant LionDocumento5 pagineShort Communication Screening of Quorum Quenching Activity of Bacteria Isolated From Ant LionRidwansyahNessuna valutazione finora
- Epigenetic Regulation in PlantsDocumento33 pagineEpigenetic Regulation in PlantsNina ReisNessuna valutazione finora
- Open Veterinary Journal, (2021), Vol. 11 (1) : 112-120: ISSN: 2226-4485 (Print) ISSN: 2218-6050 (Online) DoiDocumento9 pagineOpen Veterinary Journal, (2021), Vol. 11 (1) : 112-120: ISSN: 2226-4485 (Print) ISSN: 2218-6050 (Online) DoiVitoria de Sousa MunizNessuna valutazione finora
- 1635 (2005) Paul B. Eckburg: Science Et AlDocumento5 pagine1635 (2005) Paul B. Eckburg: Science Et AlArizha IrmaNessuna valutazione finora
- Hypersensitive Response in PlantsDocumento18 pagineHypersensitive Response in PlantsMehak MattooNessuna valutazione finora
- Possibe Association Beteween Mutldrug Resistenace and Isolate Origin With Som Virulence Factors of E Coli Strains Isolated From Infant FeacesDocumento11 paginePossibe Association Beteween Mutldrug Resistenace and Isolate Origin With Som Virulence Factors of E Coli Strains Isolated From Infant FeacesCarlos VazquezNessuna valutazione finora
- Zea Mays Spodoptera ExiguaDocumento9 pagineZea Mays Spodoptera ExiguaAimer GuzmanNessuna valutazione finora
- Arabidopsis Pathology Breathes New Life Into The Necrotrophs-Vs.-Biotrophs Classification of Fungal PathogensDocumento6 pagineArabidopsis Pathology Breathes New Life Into The Necrotrophs-Vs.-Biotrophs Classification of Fungal Pathogensyash1239Nessuna valutazione finora
- Mpmi 05 23 0073 HHDocumento8 pagineMpmi 05 23 0073 HHRoden Carlo LizardoNessuna valutazione finora
- Ige2011 PDFDocumento9 pagineIge2011 PDFAkash AroraNessuna valutazione finora
- Full PDFDocumento38 pagineFull PDFChucho RosasNessuna valutazione finora
- Molecular and Cell Biology of Arbuscular Mycorrhizal SymbiosisDocumento13 pagineMolecular and Cell Biology of Arbuscular Mycorrhizal SymbiosisGiorgio PredelliNessuna valutazione finora
- The WRKY Transcription Factor Family in Model Plants and CropsDocumento26 pagineThe WRKY Transcription Factor Family in Model Plants and CropsAndre SanchezNessuna valutazione finora
- Agent Biology-Endophytic Fungi As Biocontrol Agents of Theobroma Cacao PathogensDocumento11 pagineAgent Biology-Endophytic Fungi As Biocontrol Agents of Theobroma Cacao Pathogensglenzi fizulmiNessuna valutazione finora
- Ecology of Bacterial Endophytes in Sustainable Agriculture: Pablo R. Hardoim, Riitta Nissinen, and Jan D. Van ElsasDocumento32 pagineEcology of Bacterial Endophytes in Sustainable Agriculture: Pablo R. Hardoim, Riitta Nissinen, and Jan D. Van ElsasGregorio AroneNessuna valutazione finora
- Biological Control: Les Shipp, Jean Pierre Kapongo, Hong-Hyun Park, Peter KevanDocumento8 pagineBiological Control: Les Shipp, Jean Pierre Kapongo, Hong-Hyun Park, Peter KevanMariana AriasNessuna valutazione finora
- Infection, Genetics and EvolutionDocumento9 pagineInfection, Genetics and EvolutionAri Syuhada PutraNessuna valutazione finora
- Articulo Cientifico Estudio Comparativo de PromotoresDocumento8 pagineArticulo Cientifico Estudio Comparativo de PromotoresNINA GABRIELA SIVIZACA FLORESNessuna valutazione finora
- Update On Root Exudation and Rhizosphere BiologyDocumento8 pagineUpdate On Root Exudation and Rhizosphere BiologyNewenkeluNessuna valutazione finora
- 1Documento34 pagine1ahmredNessuna valutazione finora
- Phage Therapy: The Escherichia Coli Experience: ReviewDocumento8 paginePhage Therapy: The Escherichia Coli Experience: ReviewAbhay Kumar MohiniNessuna valutazione finora
- Rolland2016 Article QuorumSensingAndQuorumQuenchinDocumento11 pagineRolland2016 Article QuorumSensingAndQuorumQuenchinlunitajoaNessuna valutazione finora
- Stable Isotope Probing To Investigate Microbial Function in SoilDocumento23 pagineStable Isotope Probing To Investigate Microbial Function in SoilSharmin AktarNessuna valutazione finora
- 3156-2413-2-PB 2.50.26 PM 6.18.51 PMDocumento8 pagine3156-2413-2-PB 2.50.26 PM 6.18.51 PMSaiful AbdulNessuna valutazione finora
- Heritability of Oral Microbial Species in Caries-Active and Caries-FreeDocumento13 pagineHeritability of Oral Microbial Species in Caries-Active and Caries-FreeJing XueNessuna valutazione finora
- Quorum Sensing and Indole-3-Acetic Acid Degradation Play A Role in Colonization and Plant Growth Promotion of Arabidopsis Thaliana by Burkholderia Phytofirmans PSJNDocumento9 pagineQuorum Sensing and Indole-3-Acetic Acid Degradation Play A Role in Colonization and Plant Growth Promotion of Arabidopsis Thaliana by Burkholderia Phytofirmans PSJNFrancisca MartinichNessuna valutazione finora
- Dinámicas de La Biodiversidad Marina en Arrecifes de CoralDocumento7 pagineDinámicas de La Biodiversidad Marina en Arrecifes de CoralSutanejo LopezNessuna valutazione finora
- Gardner and Driks-Con Trol Biol-BacillusDocumento1 paginaGardner and Driks-Con Trol Biol-BacillusRenan Saldaña RochaNessuna valutazione finora
- Host Range Diversification Within The Incp-1 Plasmid GroupDocumento13 pagineHost Range Diversification Within The Incp-1 Plasmid GroupTiga TujuhNessuna valutazione finora
- tmpCB3A TMPDocumento4 paginetmpCB3A TMPFrontiersNessuna valutazione finora
- Fcimb 02 00090Documento7 pagineFcimb 02 00090bonniNessuna valutazione finora
- Augustinosetal 2014 RcerasiWolbachiaMicrosatsDocumento20 pagineAugustinosetal 2014 RcerasiWolbachiaMicrosatsperivou agoulaNessuna valutazione finora
- Agri Paper 4Documento12 pagineAgri Paper 4VikasNessuna valutazione finora
- Full TextDocumento10 pagineFull TextyayacandelaNessuna valutazione finora
- Science 2011 Avinoam 589 92Documento5 pagineScience 2011 Avinoam 589 92Xiomi QuirogaNessuna valutazione finora
- Cold Ham 2013Documento9 pagineCold Ham 2013Omar AguilarNessuna valutazione finora
- Intestinal Parasite Infections and Fecal Steroid Levels in Wild ChimpanzeesDocumento5 pagineIntestinal Parasite Infections and Fecal Steroid Levels in Wild ChimpanzeesDavi FragosoNessuna valutazione finora
- Discussion - Final OoDocumento3 pagineDiscussion - Final OofupaNessuna valutazione finora
- 1 s2.0 0732889391900114 Main PDFDocumento3 pagine1 s2.0 0732889391900114 Main PDFFlorencia IrenaNessuna valutazione finora
- Bioluminescence in Aquatic and Terrestrial Organisms Elicited Through Various Kinds of StimulationDocumento28 pagineBioluminescence in Aquatic and Terrestrial Organisms Elicited Through Various Kinds of StimulationHafizam YazidNessuna valutazione finora
- Won 2003Documento14 pagineWon 2003Roberto C. BárcenasNessuna valutazione finora
- The Prokaryotic Species 2020Documento29 pagineThe Prokaryotic Species 2020خديجة بيوNessuna valutazione finora
- AggloDocumento8 pagineAggloTha Joon Lautner MatsuyamaNessuna valutazione finora
- Fig 1. Diversity of Embosymbionts in Ants, Environment and Medicinal PlantsDocumento3 pagineFig 1. Diversity of Embosymbionts in Ants, Environment and Medicinal PlantsfupaNessuna valutazione finora
- Chou Et Al, 2015Documento16 pagineChou Et Al, 2015Juan RamirezNessuna valutazione finora
- Phylogenetic Evaluation of Species Nomenclature Of: Pestalotiopsis in Relation To Host AssociationDocumento17 paginePhylogenetic Evaluation of Species Nomenclature Of: Pestalotiopsis in Relation To Host AssociationAWARARAHINessuna valutazione finora
- Cox1 AgaricomycetesDocumento13 pagineCox1 AgaricomycetesLuane PortelaNessuna valutazione finora
- American Society of Plant Biologists (ASPB) The Plant CellDocumento6 pagineAmerican Society of Plant Biologists (ASPB) The Plant CellvodounnouNessuna valutazione finora
- Activation Tagging in Arabidopsis: Breakthrough TechnologiesDocumento11 pagineActivation Tagging in Arabidopsis: Breakthrough TechnologiesSujoy DebNessuna valutazione finora
- Genetics and Biotechnology of BacilliDa EverandGenetics and Biotechnology of BacilliMark M. ZukowskiNessuna valutazione finora
- Inspection and Test Plan:: PLG-SA-200-MR-MC-0005Documento4 pagineInspection and Test Plan:: PLG-SA-200-MR-MC-0005Gala BanneringgiNessuna valutazione finora
- Home RemediesDocumento18 pagineHome RemediesVardhman JainNessuna valutazione finora
- Leaves:: Form and StructureDocumento37 pagineLeaves:: Form and StructureJane ManalusNessuna valutazione finora
- Rhipsalis Baccifera: (3) (Note 1)Documento1 paginaRhipsalis Baccifera: (3) (Note 1)Tommy4Nessuna valutazione finora
- DB DipsaDocumento22 pagineDB DipsaElsa MarquezNessuna valutazione finora
- Shear and Compressive Strength Parallel To Grain of Mango Mangifera Indica at Two Moisture Regimes in A Humid EnvironmentDocumento6 pagineShear and Compressive Strength Parallel To Grain of Mango Mangifera Indica at Two Moisture Regimes in A Humid EnvironmentCrystian Kobee EmpeynadoNessuna valutazione finora
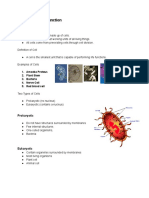
- Cell Structure FunctionDocumento4 pagineCell Structure FunctionLindley PareñoNessuna valutazione finora
- #34 Revision Sheet Plant Growth and DevelopmentDocumento4 pagine#34 Revision Sheet Plant Growth and DevelopmentAmudala HemashviniNessuna valutazione finora
- The Patagonian HerpetofaunaDocumento32 pagineThe Patagonian HerpetofaunaFernanda Alveal RiquelmeNessuna valutazione finora
- Rainbow Trout CultureDocumento6 pagineRainbow Trout Cultureluis ruperto floresNessuna valutazione finora
- Percent and PercentagesDocumento59 paginePercent and PercentagesJenn sayongNessuna valutazione finora
- MEDICINAL DIVERSITY AND ETHNOBOTANICAL USAGE OF ASTERACIOUS PLANTS OF GORAKHPUR DISTRICT, UTTAR PRADESH Srivastava C, Pande S* Department of Botany, Mahatma Gandhi P.G. College, Gorakhpur-273001, U.P., India Corresponding author: - Dr. Shail Pande, Associate Professor, Head, Dept. of Botany, Mahatma Gandhi P.G. College, Gorakhpur, U.P., India ABSTRACT The Asteraceae family is the largest family of flowering plants, with over 1600 genera and 23000 species are distributed throughout the world and occupy a wide range of habitat. In India, this family is represented by 900 species under 167 genera. Present study deals with an account of 17 medicinally important species of Asteracious plants found within present political boundaries of Gorakhpur District. The study highlights the botanical names, common names, habit, flowering and fruiting period and medicinal uses of Asteracious plants. Some important medicinal plants of this family are Ageratum conyzoides, Blumea lacera, CalenDocumento8 pagineMEDICINAL DIVERSITY AND ETHNOBOTANICAL USAGE OF ASTERACIOUS PLANTS OF GORAKHPUR DISTRICT, UTTAR PRADESH Srivastava C, Pande S* Department of Botany, Mahatma Gandhi P.G. College, Gorakhpur-273001, U.P., India Corresponding author: - Dr. Shail Pande, Associate Professor, Head, Dept. of Botany, Mahatma Gandhi P.G. College, Gorakhpur, U.P., India ABSTRACT The Asteraceae family is the largest family of flowering plants, with over 1600 genera and 23000 species are distributed throughout the world and occupy a wide range of habitat. In India, this family is represented by 900 species under 167 genera. Present study deals with an account of 17 medicinally important species of Asteracious plants found within present political boundaries of Gorakhpur District. The study highlights the botanical names, common names, habit, flowering and fruiting period and medicinal uses of Asteracious plants. Some important medicinal plants of this family are Ageratum conyzoides, Blumea lacera, CalenEarthjournal PublisherNessuna valutazione finora
- Activated Carbon From Lignocellulosics Precursors A Review of The Synthesis Methods Characterization Techniques and ApplicationsDocumento22 pagineActivated Carbon From Lignocellulosics Precursors A Review of The Synthesis Methods Characterization Techniques and ApplicationsTony OcampoNessuna valutazione finora
- 1943 7714 Article p125Documento6 pagine1943 7714 Article p125Murat AquaNessuna valutazione finora
- Roman & Byzantine LandscapeDocumento2 pagineRoman & Byzantine Landscapeaishwarya903Nessuna valutazione finora
- 01 Important Crops of The PhlippinesDocumento5 pagine01 Important Crops of The PhlippinesLaurenz James Coronado DeMattaNessuna valutazione finora
- Identifikasi Morfologi Dan Anatomi Tipe StomataDocumento9 pagineIdentifikasi Morfologi Dan Anatomi Tipe StomataDella AndreanaNessuna valutazione finora
- ARTIKELDocumento10 pagineARTIKELAn'an NuryantiNessuna valutazione finora
- BACUBACDocumento45 pagineBACUBACJamie LiwanagNessuna valutazione finora
- Biology WaecDocumento37 pagineBiology WaecPascal JohnsonNessuna valutazione finora