Potrebbero piacerti anche
- Hodgson Que Son Las Instituciones EspDocumento37 pagineHodgson Que Son Las Instituciones EspNatiResler100% (1)
- Introduccion A La Cuestion de La PsicosisDocumento15 pagineIntroduccion A La Cuestion de La PsicosisNatiReslerNessuna valutazione finora
- Antropologia Del Cuerpo y ModernidadDocumento10 pagineAntropologia Del Cuerpo y ModernidadNatiReslerNessuna valutazione finora
- Clase Ix La Metafora PaternaDocumento8 pagineClase Ix La Metafora PaternaNatiReslerNessuna valutazione finora
- Carl Gustav JungDocumento6 pagineCarl Gustav JungNatiResler100% (1)
- Repaso Tema 2Documento8 pagineRepaso Tema 2RaquelNessuna valutazione finora
- Formulario Nutricion y Dietetica en El Adulto 2020Documento17 pagineFormulario Nutricion y Dietetica en El Adulto 2020Monica Nayeli Carreon Salazar100% (2)
- Este Si 8uso Del Nopal para La Obesidad - DocxvDocumento14 pagineEste Si 8uso Del Nopal para La Obesidad - DocxvGustavo RamirezNessuna valutazione finora
- Avenas Con Frutas Exóticas-Producto PromocionDocumento9 pagineAvenas Con Frutas Exóticas-Producto PromocionMelissaNessuna valutazione finora
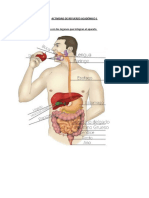
- ACTIVIDAD DE REFUERZO ACADEMICO 1 para 2CBDocumento3 pagineACTIVIDAD DE REFUERZO ACADEMICO 1 para 2CBariferr2008Nessuna valutazione finora
- Mini Lecturas Comprensivas Infantil y Primaria Temática La Prehistoria PDFDocumento5 pagineMini Lecturas Comprensivas Infantil y Primaria Temática La Prehistoria PDFYolandaNessuna valutazione finora
- Dossier-Eneagrama 2015 Online2Documento4 pagineDossier-Eneagrama 2015 Online2Mari VONessuna valutazione finora
- E-05 Ficha Tecnica Leche Uht Entera V03Documento3 pagineE-05 Ficha Tecnica Leche Uht Entera V03WALTER GUZMAN GOMEZNessuna valutazione finora
- 00 Italcol Brochure Materias Primas BajaDocumento9 pagine00 Italcol Brochure Materias Primas BajaDiego RodríguezNessuna valutazione finora
- Aminoacidos ToxicosDocumento11 pagineAminoacidos ToxicosGabyColquePacori25% (4)
- Vitamin AsDocumento11 pagineVitamin AsGamalielNessuna valutazione finora
- PROYECTO Paletas de HieloDocumento12 paginePROYECTO Paletas de HieloJUAN PABLO HERNANDEZ PEREZNessuna valutazione finora
- TEMA 3 Nutrición y AlimentaciónDocumento4 pagineTEMA 3 Nutrición y AlimentaciónAlejandro FernandezNessuna valutazione finora
- Método de Microencapsulación de Cobertura Por Lecho FluidizadoDocumento29 pagineMétodo de Microencapsulación de Cobertura Por Lecho FluidizadoItzel De los santos GutiérrezNessuna valutazione finora
- PRACTICA8Documento6 paginePRACTICA8Tyler de los santos 21Nessuna valutazione finora
- Tamizaje Con HerramientaDocumento1 paginaTamizaje Con HerramientaGeiner Romero Zapata100% (2)
- Proyecto ProductivoDocumento5 pagineProyecto ProductivoJC HANessuna valutazione finora
- 1Documento11 pagine1Cruzito ClNessuna valutazione finora
- Diptico Cuidado de La SaludDocumento4 pagineDiptico Cuidado de La SaludGean KalinNessuna valutazione finora
- Propuesta de Plan SemanalDocumento3 paginePropuesta de Plan SemanalJUANNessuna valutazione finora
- Crema Budwig Alternativa Dra KousmineDocumento6 pagineCrema Budwig Alternativa Dra KousminePaul Peter PaulNessuna valutazione finora
- Tesis Produccion y Calidad de Leche de Vacas en Pastoreo o en EstabulacionDocumento86 pagineTesis Produccion y Calidad de Leche de Vacas en Pastoreo o en EstabulacionPatricia CordovaNessuna valutazione finora
- Fichas Grafico DescriptivaDocumento10 pagineFichas Grafico DescriptivaJechu RiveraNessuna valutazione finora
- Swipe E-Catalogo Domestico 2020Documento17 pagineSwipe E-Catalogo Domestico 2020swipe huajuapanNessuna valutazione finora
- Rien Documento Tecnico PDFDocumento42 pagineRien Documento Tecnico PDFconsuelo aguilarNessuna valutazione finora
- Cueva Lopez Milton-EconomíaDocumento10 pagineCueva Lopez Milton-EconomíaMilton Cueva LópezNessuna valutazione finora
- Enjuagues Bucales Con Carbohidratos y Su Efecto en El Rendimiento de Carreras Contrarreloj: Revisión SistemáticaDocumento8 pagineEnjuagues Bucales Con Carbohidratos y Su Efecto en El Rendimiento de Carreras Contrarreloj: Revisión SistemáticaSHAYRA NORIEGA ROSSONessuna valutazione finora
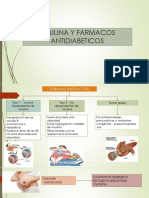
- AntidiabeticosDocumento49 pagineAntidiabeticosVíctor Manuel100% (3)
- Comportamiento Caprino PDFDocumento15 pagineComportamiento Caprino PDFJuan VelascoNessuna valutazione finora
- MORGADO111Documento9 pagineMORGADO111Paolo PerezvilcaNessuna valutazione finora
- Tus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadDa EverandTus Zonas Erroneas: Guía Para Combatir las Causas de la InfelicidadValutazione: 4.5 su 5 stelle4.5/5 (1833)
- Los Secretos De La Mente Millonaria: Domina el juego de la riquezaDa EverandLos Secretos De La Mente Millonaria: Domina el juego de la riquezaValutazione: 5 su 5 stelle5/5 (457)
- Resetea tu mente. Descubre de lo que eres capazDa EverandResetea tu mente. Descubre de lo que eres capazValutazione: 5 su 5 stelle5/5 (196)
- El Monje Que Vendio Su Ferrari: Una Fábula EspiritualDa EverandEl Monje Que Vendio Su Ferrari: Una Fábula EspiritualValutazione: 4.5 su 5 stelle4.5/5 (1699)
- Ayurveda: Descubriendo los Secretos de la Curación Hindú a Través de la Dieta del Ayurveda, el Yoga, la Aromaterapia y la MeditaciónDa EverandAyurveda: Descubriendo los Secretos de la Curación Hindú a Través de la Dieta del Ayurveda, el Yoga, la Aromaterapia y la MeditaciónValutazione: 4 su 5 stelle4/5 (7)
- Guías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!Da EverandGuías Espirituales: ¡Una guía para conectarte y comunicarte con tus guías espirituales!Valutazione: 4.5 su 5 stelle4.5/5 (14)
- La metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceDa EverandLa metamedicina. Cada síntoma es un mensaje: La curación a tu alcanceValutazione: 5 su 5 stelle5/5 (8)
- ¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadDa Everand¡Tómate un respiro! Mindfulness: El arte de mantener la calma en medio de la tempestadValutazione: 5 su 5 stelle5/5 (199)
- Escribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelDa EverandEscribe el guion de la vida que quieres: Manifiesta tus sueños con tan solo lápiz y papelValutazione: 5 su 5 stelle5/5 (1)
- Homo antecessor: El nacimiento de una especieDa EverandHomo antecessor: El nacimiento de una especieValutazione: 5 su 5 stelle5/5 (1)
- Entrenamiento Científico con pesas: Fitness InteligenteDa EverandEntrenamiento Científico con pesas: Fitness InteligenteValutazione: 5 su 5 stelle5/5 (3)