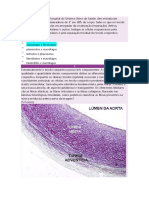
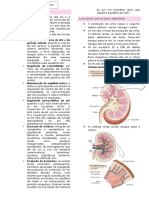
Potrebbero piacerti anche
- Os 5 elementos da medicina tradicional chinesaDocumento1 paginaOs 5 elementos da medicina tradicional chinesaAnonymous OdfoTPUNessuna valutazione finora
- Restauração do organismo humano em númerosDocumento107 pagineRestauração do organismo humano em númerosLeila Martins100% (6)
- Osteopatia GargantaDocumento15 pagineOsteopatia GargantaCaius CunhaNessuna valutazione finora
- PATOLOGIA 04 - Inflamação Crônica - MED RESUMOS - Arlindo NettoDocumento7 paginePATOLOGIA 04 - Inflamação Crônica - MED RESUMOS - Arlindo NettoFernando Zucuni FurlanNessuna valutazione finora
- Estudo Dirigido de Aulas Práticas - Fag ParanáDocumento31 pagineEstudo Dirigido de Aulas Práticas - Fag ParanáVanessaDoEsa100% (1)
- Book 21235Documento248 pagineBook 21235Jayllon HenriqueNessuna valutazione finora
- Atividade Sub - Bedu - Fisiologia Geral e Do Movimento - 54-2022Documento7 pagineAtividade Sub - Bedu - Fisiologia Geral e Do Movimento - 54-2022Cavalini Assessoria Acadêmica0% (2)
- PATOLOGIA 02 - Lesão e Morte Celular - MED RESUMOS - Arlindo Netto PDFDocumento7 paginePATOLOGIA 02 - Lesão e Morte Celular - MED RESUMOS - Arlindo Netto PDFGabriela Grijp100% (1)
- PATOLOGIA 10 - Fisiopatogênese Da AteroscleroseDocumento6 paginePATOLOGIA 10 - Fisiopatogênese Da AterosclerosePaulo SpíndolaNessuna valutazione finora
- Filo Arthropoda - Características e TaxonomiaDocumento34 pagineFilo Arthropoda - Características e TaxonomiaDavid Da Silva ConstantinoNessuna valutazione finora
- Clinica Ampliada CartilhaDocumento68 pagineClinica Ampliada CartilhaMichelly MoreiraNessuna valutazione finora
- Resumão Pra Felicidade - 1 ProvaDocumento7 pagineResumão Pra Felicidade - 1 ProvaPaulo SpíndolaNessuna valutazione finora
- Anamnes eDocumento9 pagineAnamnes ePaulo SpíndolaNessuna valutazione finora
- Adm Questões NovasDocumento23 pagineAdm Questões NovasPaulo SpíndolaNessuna valutazione finora
- ArtrópodesDocumento35 pagineArtrópodesPaulo SpíndolaNessuna valutazione finora
- Humanização da saúde no BrasilDocumento22 pagineHumanização da saúde no BrasilPaulo SpíndolaNessuna valutazione finora
- Portaria204 17 Fevereiro 2016Documento7 paginePortaria204 17 Fevereiro 2016Luis Carlos RosaNessuna valutazione finora
- 3 - Desenvolvimento - PAULODocumento3 pagine3 - Desenvolvimento - PAULOPaulo SpíndolaNessuna valutazione finora
- PATOLOGIA 07 - Doenças Infecciosas - MED RESUMOS - Arlindo NettoDocumento10 paginePATOLOGIA 07 - Doenças Infecciosas - MED RESUMOS - Arlindo NettoPaulo SpíndolaNessuna valutazione finora
- Acolhimento No BrasilDocumento11 pagineAcolhimento No BrasilPaulo SpíndolaNessuna valutazione finora
- ApoptoseDocumento9 pagineApoptoseGetúlio Bomfim100% (1)
- Humanização Na Atenção Basica - Acolhimento e Clinica Ampliada - Revisado PAULO.Documento6 pagineHumanização Na Atenção Basica - Acolhimento e Clinica Ampliada - Revisado PAULO.Paulo SpíndolaNessuna valutazione finora
- Neoplasia Patologia MédicaDocumento13 pagineNeoplasia Patologia MédicaJúlia Oliveira de CarvalhoNessuna valutazione finora
- PATOLOGIA 09 - Pigmentação PatológicaDocumento6 paginePATOLOGIA 09 - Pigmentação PatológicaPaulo SpíndolaNessuna valutazione finora
- 1 Código Penal Militar EAP Crime Militar 15 1 15 PDFDocumento8 pagine1 Código Penal Militar EAP Crime Militar 15 1 15 PDFPaulo SpíndolaNessuna valutazione finora
- Paulo Tadeu Comentarios Aos Arts 01 A 37 Do Codigo Penal Militar EbookDocumento59 paginePaulo Tadeu Comentarios Aos Arts 01 A 37 Do Codigo Penal Militar EbooksaviosquasherNessuna valutazione finora
- M 2 PM Manual de Policiamento de Transito UrbanoDocumento31 pagineM 2 PM Manual de Policiamento de Transito UrbanoPaulo SpíndolaNessuna valutazione finora
- Freqüência de Doenças ReumáticasDocumento8 pagineFreqüência de Doenças ReumáticasPaulo SpíndolaNessuna valutazione finora
- Atlas de ParasitoDocumento4 pagineAtlas de ParasitoPaulo SpíndolaNessuna valutazione finora
- Inflamação Na Doença Renal Crônica Papel de CitocinasDocumento14 pagineInflamação Na Doença Renal Crônica Papel de CitocinasPaulo SpíndolaNessuna valutazione finora
- Provimento 09-2011Documento7 pagineProvimento 09-2011Paulo SpíndolaNessuna valutazione finora
- Denúncia contra policiais por crimes políticosDocumento83 pagineDenúncia contra policiais por crimes políticosPaulo SpíndolaNessuna valutazione finora
- Dos Crimes de Abandono de PostoDocumento26 pagineDos Crimes de Abandono de PostoEduardo MoreiraNessuna valutazione finora
- Cronograma - MicroDocumento1 paginaCronograma - MicroPaulo SpíndolaNessuna valutazione finora
- Cronograma - MicroDocumento1 paginaCronograma - MicroPaulo SpíndolaNessuna valutazione finora
- 34 - Resumo VII Berne - Elementos Da Função RenalDocumento1 pagina34 - Resumo VII Berne - Elementos Da Função RenalPaulo SpíndolaNessuna valutazione finora
- Portaria #2488Documento28 paginePortaria #2488Paulo SpíndolaNessuna valutazione finora
- Aluna: Ana Paula Souza Gomes Biossistemas Do Corpo HumanoDocumento1 paginaAluna: Ana Paula Souza Gomes Biossistemas Do Corpo HumanoAna Paula GomesNessuna valutazione finora
- ELETROMICROGRAFIAS - ExplicadasDocumento132 pagineELETROMICROGRAFIAS - ExplicadasRomana GalleteNessuna valutazione finora
- Histologia Dos Vasos SanguineosDocumento3 pagineHistologia Dos Vasos SanguineosbeatrizNessuna valutazione finora
- Obstrução Intestinal Por Necrose Massiva de Gordura Abdominal (Lipomatose) em Uma VacaDocumento3 pagineObstrução Intestinal Por Necrose Massiva de Gordura Abdominal (Lipomatose) em Uma VacaAnselmo RodriguezNessuna valutazione finora
- Mecanismos Fisiopatológicos de Alteração Da Capacidade Funcional em CardiopatasDocumento10 pagineMecanismos Fisiopatológicos de Alteração Da Capacidade Funcional em CardiopatasAderval NetoNessuna valutazione finora
- Regulação da reprodução pelo sistema neural e neuroendócrinoDocumento2 pagineRegulação da reprodução pelo sistema neural e neuroendócrinoBruna VieiraNessuna valutazione finora
- Características tecido encontrado em Serra da CapivaraDocumento7 pagineCaracterísticas tecido encontrado em Serra da CapivaraSabrina França CardosoNessuna valutazione finora
- Anatomia CorrigidaDocumento10 pagineAnatomia Corrigidasandrobastos100% (3)
- Sistema Respiratório: Função, Estrutura e MecanismosDocumento4 pagineSistema Respiratório: Função, Estrutura e MecanismosCinthia CananNessuna valutazione finora
- Animais Tambem Têm Tipo de Sangue Igual Ao NossoDocumento5 pagineAnimais Tambem Têm Tipo de Sangue Igual Ao NossoDAVID ALEXANDRE ROSA CRUZNessuna valutazione finora
- Ficha de Leitura - PsicDocumento5 pagineFicha de Leitura - Psicsaidefelix mariaNessuna valutazione finora
- Sistema SensorialDocumento26 pagineSistema SensorialMaria Cristina CristinaNessuna valutazione finora
- Causas Das DoençasDocumento8 pagineCausas Das DoençasMalu1961Nessuna valutazione finora
- Alterações Pulpares - UnespDocumento14 pagineAlterações Pulpares - UnespTainá OnoNessuna valutazione finora
- 5 - RESUMO LINFÓCITO B (GERAÇÃO, ATIVAÇÃO e PRODUÇÃO ANTICORPOS)Documento4 pagine5 - RESUMO LINFÓCITO B (GERAÇÃO, ATIVAÇÃO e PRODUÇÃO ANTICORPOS)ImunoResumo93% (15)
- Invertebrados IDocumento12 pagineInvertebrados IGabriella Rodrigues SilvaNessuna valutazione finora
- Fisiologia CardíacaDocumento7 pagineFisiologia CardíacaAndreza LuciaNessuna valutazione finora
- SufixosDocumento4 pagineSufixosDayane FerreiraNessuna valutazione finora
- Fisiologia Do Sistema UrinárioDocumento7 pagineFisiologia Do Sistema UrinárioRafaela SoratoNessuna valutazione finora
- Afecc;oes do Aparelho Locomotor (ossos e articulac;oesDocumento41 pagineAfecc;oes do Aparelho Locomotor (ossos e articulac;oesFelipe VianaNessuna valutazione finora
- 2.1 Estruturas e Divisão Anatômica Da MedulaDocumento42 pagine2.1 Estruturas e Divisão Anatômica Da MedulacarolinamarinhoguiNessuna valutazione finora
- Sistema Olfativo ComponentesDocumento6 pagineSistema Olfativo ComponentesviniciusNessuna valutazione finora
- Aula - Controle Da Ecdise e MetamorfoseDocumento32 pagineAula - Controle Da Ecdise e MetamorfoseValdinei Junio Brito VilelaNessuna valutazione finora