Potrebbero piacerti anche
- Guia de Estudio #6 Célula Procariota y EucariotaDocumento7 pagineGuia de Estudio #6 Célula Procariota y EucariotaceceortNessuna valutazione finora
- 09 - BiosensoresDocumento60 pagine09 - BiosensoresLuis Rodríguez100% (1)
- Obtener Archivo RecursoDocumento6 pagineObtener Archivo RecursoJhon CarrionNessuna valutazione finora
- ETOGRAMADocumento10 pagineETOGRAMAjhon muesesNessuna valutazione finora
- Tesis Fitoplancton FranciscoDocumento95 pagineTesis Fitoplancton FranciscoAndrea BANessuna valutazione finora
- Reconocimiento de protozoosDocumento6 pagineReconocimiento de protozoosSergio BejaranoNessuna valutazione finora
- Practica 1 Descripcion Material de LaboratorioDocumento13 paginePractica 1 Descripcion Material de LaboratorioMarcia NisNessuna valutazione finora
- Protozoos parásitosDocumento9 pagineProtozoos parásitossheriff ChávezNessuna valutazione finora
- Metabolismo BacterianoDocumento46 pagineMetabolismo BacterianoAndres Flores PardaveNessuna valutazione finora
- Lab2 ZooplanctonDocumento14 pagineLab2 ZooplanctonCACERES ESTRADA IACOMO FERNANDO100% (1)
- Práctica 8Documento3 paginePráctica 8Stefany Asmat D'AngeloNessuna valutazione finora
- Practica 1 Anatomía Interna y Externa de Peces ÓseosDocumento4 paginePractica 1 Anatomía Interna y Externa de Peces ÓseosBetty CaSu100% (1)
- Tecnicas AvesDocumento13 pagineTecnicas AvesJean CarlosNessuna valutazione finora
- NITRILOSDocumento11 pagineNITRILOSAndres A. Rocha UsugaNessuna valutazione finora
- BacillariophytaDocumento37 pagineBacillariophytaKAREN MARGARITA PACHECO SALINAS100% (2)
- Manual Laboratorio Bio-011, Bio-010 Uce. Version 1. Enero 2018 (1)Documento119 pagineManual Laboratorio Bio-011, Bio-010 Uce. Version 1. Enero 2018 (1)Charina De la Cruz0% (1)
- Identificación de estructuras celulares vegetalesDocumento8 pagineIdentificación de estructuras celulares vegetalesPablo CruzNessuna valutazione finora
- Mioseptos y MiomerosDocumento5 pagineMioseptos y MiomerosSamy Carballido HuamanNessuna valutazione finora
- Sistema de Soporte y Locomocion en Los SeresDocumento21 pagineSistema de Soporte y Locomocion en Los SeresEver VanegasNessuna valutazione finora
- Uam ProtozoariosDocumento37 pagineUam ProtozoariosTeofagia Mariana100% (1)
- Identificacion de Copépodos y EufausidosDocumento7 pagineIdentificacion de Copépodos y EufausidosIce CreamNessuna valutazione finora
- Práctica 1 - MicroalgasDocumento10 paginePráctica 1 - MicroalgasWero Bc100% (1)
- GlicocalixDocumento12 pagineGlicocalixTeo NavarroNessuna valutazione finora
- Protozoarios 1Documento19 pagineProtozoarios 1HansPomaMolleapazaNessuna valutazione finora
- ChaetognathaDocumento3 pagineChaetognathaJussel Ofir Vázquez MárquezNessuna valutazione finora
- PRÁCTICA No 1-BIOSEGURIDAD-BB - I 2020Documento5 paginePRÁCTICA No 1-BIOSEGURIDAD-BB - I 2020sergio amayaNessuna valutazione finora
- Articulo Cientifico Invertebrados AtpDocumento6 pagineArticulo Cientifico Invertebrados AtpmichaelNessuna valutazione finora
- Práctica 11 - Plasmólisis y TurgenciaDocumento3 paginePráctica 11 - Plasmólisis y TurgenciaMónica CantúNessuna valutazione finora
- Practica N 8 Fenomenos Fisicos de La Celula Difusion OsmosisDocumento13 paginePractica N 8 Fenomenos Fisicos de La Celula Difusion OsmosisJesus E. Cabezudo CalderonNessuna valutazione finora
- Guía Teórica Práctica Poriferos 5ºbmayo 2010nombreDocumento5 pagineGuía Teórica Práctica Poriferos 5ºbmayo 2010nombreOscar100% (1)
- Algas IntroduccionDocumento7 pagineAlgas IntroduccionEdu Cristian Calle MorochoNessuna valutazione finora
- Microorganismos de Agua DulceDocumento3 pagineMicroorganismos de Agua DulceKAREN RIBERONessuna valutazione finora
- Características de Las Algas. Cuadro ComparativoDocumento3 pagineCaracterísticas de Las Algas. Cuadro ComparativoJazmín CalixtoNessuna valutazione finora
- Manejo y Uso Del Microscopio Óptico CompuestoDocumento25 pagineManejo y Uso Del Microscopio Óptico Compuestoperito29Nessuna valutazione finora
- Biometria de AvesDocumento1 paginaBiometria de AvesNicolas WilliamNessuna valutazione finora
- Histologia de Mucosa Olfatoria y Corpusculos GustativosDocumento3 pagineHistologia de Mucosa Olfatoria y Corpusculos GustativosDiana OlivaresNessuna valutazione finora
- Trabajo Rhizopodos ArlethDocumento11 pagineTrabajo Rhizopodos ArlethArleth ZeballosNessuna valutazione finora
- Practica Sistema EndócrinoDocumento4 paginePractica Sistema EndócrinoLili MïrandaNessuna valutazione finora
- Introducción Al Manejo de DrosophilaDocumento8 pagineIntroducción Al Manejo de DrosophilaAndrea BonillaNessuna valutazione finora
- Practica No 6 Reptiles IDocumento18 paginePractica No 6 Reptiles Iaea123123123Nessuna valutazione finora
- ADN D La FrezaDocumento1 paginaADN D La FrezaJenath RamosNessuna valutazione finora
- Práctica de Laboratorio Nro 2 Phylum PoriferaDocumento13 paginePráctica de Laboratorio Nro 2 Phylum PoriferaDaniela OrtegaNessuna valutazione finora
- Características de Los AnfibiosDocumento10 pagineCaracterísticas de Los AnfibiosGemulinNessuna valutazione finora
- Agua y Sales MineralesDocumento25 pagineAgua y Sales MineralesVICENTE100% (2)
- Columna Winogradsky: Microorganismos y zonasDocumento17 pagineColumna Winogradsky: Microorganismos y zonasLiloMontesinoNessuna valutazione finora
- ProtozoosAcuaticosDocumento4 pagineProtozoosAcuaticosjosebioNessuna valutazione finora
- Alcoholes Fenoles Eteres y TiolesDocumento49 pagineAlcoholes Fenoles Eteres y TiolesWily Pesantez100% (1)
- Poríferos: morfología y características de las esponjasDocumento11 paginePoríferos: morfología y características de las esponjasJuan Sebastián RivasNessuna valutazione finora
- Laboratorio No. 2 MicrosDocumento2 pagineLaboratorio No. 2 MicrosNuñez BrayanNessuna valutazione finora
- MACROINVERTEBRADOSDocumento26 pagineMACROINVERTEBRADOSWilder Doneth Oblitas AbantoNessuna valutazione finora
- Rutas FermentDocumento32 pagineRutas FermentAna Isabel Díaz0% (1)
- Informe Practica 4Documento8 pagineInforme Practica 4Alexandra MoralesNessuna valutazione finora
- Informe de Zoología 5 (Identificación de Un Coral (Pseudodiploria Strigosa) ) .Documento4 pagineInforme de Zoología 5 (Identificación de Un Coral (Pseudodiploria Strigosa) ) .Lyons Andrew BerrnatNessuna valutazione finora
- Guia Adhesion Celular 2011Documento4 pagineGuia Adhesion Celular 2011Clarys DiazNessuna valutazione finora
- Esquema de Sesión de Aprendizaje EvelynDocumento6 pagineEsquema de Sesión de Aprendizaje EvelynAdriano Santisteban RivasNessuna valutazione finora
- Fecundacion y EmbarazoDocumento33 pagineFecundacion y EmbarazoCamila Castillo100% (1)
- Bioplásticos, en Pro Del Ambiente - Descubre Fundación UNAMDocumento9 pagineBioplásticos, en Pro Del Ambiente - Descubre Fundación UNAMCarlos Felipe Peña MalacaraNessuna valutazione finora
- ProteinasDocumento4 pagineProteinashata_66Nessuna valutazione finora
- Libro Universidad NacionalDocumento8 pagineLibro Universidad NacionalCarolina Ortiz PinedaNessuna valutazione finora
- Boletín - Aborda Investigador de UAEH Las Nuevas MasculinidadesDocumento3 pagineBoletín - Aborda Investigador de UAEH Las Nuevas MasculinidadesmejiareyescarlosNessuna valutazione finora
- La Ganadería Bovina en TabascoDocumento21 pagineLa Ganadería Bovina en TabascoRaul KimNessuna valutazione finora
- Fisiología Reproductiva y Diferencias Reproductivas Entre El Ganado Europeo y Cebú.Documento5 pagineFisiología Reproductiva y Diferencias Reproductivas Entre El Ganado Europeo y Cebú.Raul KimNessuna valutazione finora
- Presentación DepredaciónDocumento20 paginePresentación DepredaciónRaul KimNessuna valutazione finora
- +efecto de Los Factores Causantes de Estres en Bovinos Del Trópico Húmedo-De DiosDocumento6 pagine+efecto de Los Factores Causantes de Estres en Bovinos Del Trópico Húmedo-De DiosRaul KimNessuna valutazione finora
- Ecofisiología de Los Bovinos en Sistemas de Producción Del Trópico Húmedo-De Dios.Documento409 pagineEcofisiología de Los Bovinos en Sistemas de Producción Del Trópico Húmedo-De Dios.Raul Kim100% (1)
- Presentación DepredaciónDocumento20 paginePresentación DepredaciónRaul KimNessuna valutazione finora
- Elementos Del Clima Que Afectan La Ganaderia 2019-UODocumento53 pagineElementos Del Clima Que Afectan La Ganaderia 2019-UORaul Kim100% (1)
- Industria Del Huevo en MéxicoDocumento6 pagineIndustria Del Huevo en MéxicoRaul KimNessuna valutazione finora
- Analisis de Crecimiento Del Pasto Maralfalfa en Clima Cálido Subhúmedo.Documento14 pagineAnalisis de Crecimiento Del Pasto Maralfalfa en Clima Cálido Subhúmedo.Raul KimNessuna valutazione finora
- Creep FeedingDocumento10 pagineCreep FeedingRaul KimNessuna valutazione finora
- Presentacion-Músculos de Cortes BovinosDocumento20 paginePresentacion-Músculos de Cortes BovinosRaul KimNessuna valutazione finora
- IntelliBond C-Revista Mundo Ganadero 2012.Documento68 pagineIntelliBond C-Revista Mundo Ganadero 2012.Raul KimNessuna valutazione finora
- Presentacion-Raul Kim - 2Documento35 paginePresentacion-Raul Kim - 2Raul KimNessuna valutazione finora
- MURRAY-Hormonas Exógenas en Carne de Pollo-Creencias Populares y EvidenciasDocumento14 pagineMURRAY-Hormonas Exógenas en Carne de Pollo-Creencias Populares y EvidenciasJared Nun Barrenechea BacaNessuna valutazione finora
- Presentacion-Raul Kim - 2Documento35 paginePresentacion-Raul Kim - 2Raul KimNessuna valutazione finora
- Cercas Vivas de Madero Negro.Documento7 pagineCercas Vivas de Madero Negro.Raul KimNessuna valutazione finora
- Presentacion-Raul Kim - 2Documento35 paginePresentacion-Raul Kim - 2Raul KimNessuna valutazione finora
- Dinamica de Crecimiento de La Maralfalfa en Condiciones de La Amazonia Ecuatoriana.Documento5 pagineDinamica de Crecimiento de La Maralfalfa en Condiciones de La Amazonia Ecuatoriana.Raul KimNessuna valutazione finora
- Néstor Valencia - La Gallina Criolla ColombianaDocumento64 pagineNéstor Valencia - La Gallina Criolla ColombianaAndrés Cuellar Saenz100% (1)
- Calidad Nutricional Del Pasto Maralfalfa (Pennisetum SP) Cosechado A Dos Edades de RebroteDocumento14 pagineCalidad Nutricional Del Pasto Maralfalfa (Pennisetum SP) Cosechado A Dos Edades de RebroteRaul KimNessuna valutazione finora
- Presentacion Diplomado de Vacunologia UNAM FMVZ.Documento89 paginePresentacion Diplomado de Vacunologia UNAM FMVZ.Raul Kim100% (1)
- Aspectos Claves en El Transporte de Pollos-Selecciones AvicolasDocumento4 pagineAspectos Claves en El Transporte de Pollos-Selecciones AvicolasRaul KimNessuna valutazione finora
- Densidad de GallinasDocumento30 pagineDensidad de GallinasbelindaNessuna valutazione finora
- Tesis-Tasas de Gestacion en Novillonas Peripuberes Despues de Un Tratamiento Sincronizador de La Ovulacion Con Norgestomet (Crestar) y GNRH y PGF2aDocumento36 pagineTesis-Tasas de Gestacion en Novillonas Peripuberes Despues de Un Tratamiento Sincronizador de La Ovulacion Con Norgestomet (Crestar) y GNRH y PGF2aRaul KimNessuna valutazione finora
- #Dilepididosis Amoebotaenia, Estudio de Caso.Documento1 pagina#Dilepididosis Amoebotaenia, Estudio de Caso.Raul KimNessuna valutazione finora
- #Prevalencia y Carga Parasitaria de Helmintos Gastrointestinales en Gallinas de TraspatioDocumento5 pagine#Prevalencia y Carga Parasitaria de Helmintos Gastrointestinales en Gallinas de TraspatioRaul KimNessuna valutazione finora
- Ectoparásitos de aves de corral en el sur de MéxicoDocumento5 pagineEctoparásitos de aves de corral en el sur de MéxicoRaul KimNessuna valutazione finora
- Bienestar AvicolaDocumento8 pagineBienestar AvicolaRaul KimNessuna valutazione finora
- Efecto de La Altura de Los Comederos Sobre El Largo Del Tarso, Pigmentacion y Parametros Productivos en Pollo de EngordaDocumento7 pagineEfecto de La Altura de Los Comederos Sobre El Largo Del Tarso, Pigmentacion y Parametros Productivos en Pollo de EngordaRaul KimNessuna valutazione finora
- +en Defensa de La Tesis y en Contra de La Universidad Como Expendedora de Títulos - Crítica ÁcidaDocumento5 pagine+en Defensa de La Tesis y en Contra de La Universidad Como Expendedora de Títulos - Crítica ÁcidaRaul KimNessuna valutazione finora
- Qué Dice El CIE 10 Sobre El AlzheimerDocumento2 pagineQué Dice El CIE 10 Sobre El AlzheimeresperanzaNessuna valutazione finora
- Consecuencias Del Consumo de DrogasDocumento5 pagineConsecuencias Del Consumo de DrogasLa FliaNessuna valutazione finora
- Abordaje KawaseDocumento4 pagineAbordaje KawaseAndy AgrelaNessuna valutazione finora
- Repaso Examen 2 de PsicologiaDocumento12 pagineRepaso Examen 2 de PsicologiafaustoNessuna valutazione finora
- Empatía en Trastorno Mental Grave PDFDocumento8 pagineEmpatía en Trastorno Mental Grave PDFRutNessuna valutazione finora
- La psicología, la cognición y el cerebroDocumento3 pagineLa psicología, la cognición y el cerebroMETRO PAC100% (1)
- Tesis Maestria - Análisis Neuropsicológico de La Atención, Memoria y Funciones Ejecutivas en Niños Con TdahDocumento89 pagineTesis Maestria - Análisis Neuropsicológico de La Atención, Memoria y Funciones Ejecutivas en Niños Con TdahTURK2YNessuna valutazione finora
- Médula Espinal: Morfología ExternaDocumento37 pagineMédula Espinal: Morfología ExternaInesNessuna valutazione finora
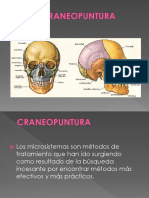
- Craneopuntura: microsistema terapéutico basado en la estimulación de zonas cranealesDocumento55 pagineCraneopuntura: microsistema terapéutico basado en la estimulación de zonas cranealesGuadalupe Salas Martinez88% (8)
- Lenguaje y CerebroDocumento7 pagineLenguaje y CerebroCeledith PiñaNessuna valutazione finora
- Tarea Nro 2 Ao 2 MFH II Semana 2 5 Al 10 de Julio de 2021Documento4 pagineTarea Nro 2 Ao 2 MFH II Semana 2 5 Al 10 de Julio de 2021Prof. Jose MurilloNessuna valutazione finora
- El Sistema Nervios1Documento4 pagineEl Sistema Nervios1DayiiTANessuna valutazione finora
- Dis LaliaDocumento2 pagineDis LaliaJorge Félix González AcostaNessuna valutazione finora
- Tarea 1 Como El Cerebro Da Lugar A La MenteDocumento6 pagineTarea 1 Como El Cerebro Da Lugar A La MenteVictoria Perez100% (1)
- 3 Suicidio TeoríasDocumento28 pagine3 Suicidio TeoríasmariloNessuna valutazione finora
- Atención y Memoria (Neuroeducación)Documento10 pagineAtención y Memoria (Neuroeducación)Edison GonzalezNessuna valutazione finora
- Histologia - Cuestionario GrupalDocumento44 pagineHistologia - Cuestionario GrupalJosé AraúzNessuna valutazione finora
- Blank DiagramDocumento1 paginaBlank DiagramIsaac Jehu Vazquez Ayala100% (1)
- Terapia Ocupacional en Sordos - Doc Versión 1Documento3 pagineTerapia Ocupacional en Sordos - Doc Versión 1Fedora Margarita Santander CeronNessuna valutazione finora
- Mapa Conceptual #2Documento1 paginaMapa Conceptual #2Luz Dary Perez contrerasNessuna valutazione finora
- Qué Es La NeurocienciaDocumento40 pagineQué Es La NeurocienciaesteralmeydaNessuna valutazione finora
- Mapa Conceptual, Tipos de NeuronaDocumento2 pagineMapa Conceptual, Tipos de NeuronaMirê IdkNessuna valutazione finora
- Análisis Articulo JazminDocumento3 pagineAnálisis Articulo JazminJazmin Garcia BergesNessuna valutazione finora
- Ensayo de Gerontologia-1Documento9 pagineEnsayo de Gerontologia-1Lizz PeñaNessuna valutazione finora
- Intervención de Terapia Ocupacional en La Rehabilitación Neurológica Subaguda Luego de Un Accidente Cerebro Vascular - Reporte de Caso ClínicoDocumento3 pagineIntervención de Terapia Ocupacional en La Rehabilitación Neurológica Subaguda Luego de Un Accidente Cerebro Vascular - Reporte de Caso ClínicoMarcia Carolina PérezNessuna valutazione finora
- Hiperalgesia Inducida Por OpiáceosDocumento7 pagineHiperalgesia Inducida Por OpiáceossanjuandediosanestesiaNessuna valutazione finora
- Abordaje Terapeutico en PC Espastico HemiplejiaDocumento40 pagineAbordaje Terapeutico en PC Espastico HemiplejiaJorge Joel Salazar MirandaNessuna valutazione finora
- Ejercicios para AfasiasDocumento2 pagineEjercicios para AfasiasHELENALISON100% (2)
- Médula EspinalDocumento33 pagineMédula EspinalWillirNessuna valutazione finora
- ParkinsonDocumento10 pagineParkinsonMARIANA BALCÁZAR TORRESNessuna valutazione finora