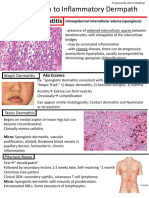
Potrebbero piacerti anche
- The Latest On Skin Photoprotection: Salvador González, MD, Manuel Fernández-Lorente, MD, Yolanda Gilaberte-Calzada, MDDocumento13 pagineThe Latest On Skin Photoprotection: Salvador González, MD, Manuel Fernández-Lorente, MD, Yolanda Gilaberte-Calzada, MDKarina Terry AmartiniNessuna valutazione finora
- Eczema and PsoriasisDocumento16 pagineEczema and PsoriasisLamy SNessuna valutazione finora
- Moisturizers The Slippery RoadDocumento9 pagineMoisturizers The Slippery RoadBERNADETH YOSTEFANY100% (1)
- N - Furfuryladenine (Kinetin) As A Potential Anti-Aging MoleculeDocumento4 pagineN - Furfuryladenine (Kinetin) As A Potential Anti-Aging MoleculeLingka EpsNessuna valutazione finora
- Acne ConglobataDocumento6 pagineAcne ConglobataClauNessuna valutazione finora
- Wet Dressings A3Documento1 paginaWet Dressings A3Camden Roberts100% (1)
- PPK Perdoski 2017 Skabies (English)Documento3 paginePPK Perdoski 2017 Skabies (English)yumizoneNessuna valutazione finora
- Multidrug Resistance EssayDocumento15 pagineMultidrug Resistance EssayShainaz Mahmood Shams AddeenNessuna valutazione finora
- British Journal of Nutrition (2008), 99, E-Suppl.Documento50 pagineBritish Journal of Nutrition (2008), 99, E-Suppl.nihadtoussounNessuna valutazione finora
- Steroid Pulse Therapies in DermatologyDocumento4 pagineSteroid Pulse Therapies in DermatologyWelly WijayantiNessuna valutazione finora
- J. Chem. Pharm. Res.,2010, 2 (1) 385-395Documento11 pagineJ. Chem. Pharm. Res.,2010, 2 (1) 385-395Irfan AijazNessuna valutazione finora
- Eczema and Nummular Dermatitis GuideDocumento91 pagineEczema and Nummular Dermatitis GuideJhost Clinton PurbaNessuna valutazione finora
- Fixed Drug EruptionDocumento13 pagineFixed Drug EruptionDian Atillah IkhwaniNessuna valutazione finora
- Referat Kandidosis Kutis - Dhira Dan A. Fari-70700120034 Dan 038Documento14 pagineReferat Kandidosis Kutis - Dhira Dan A. Fari-70700120034 Dan 038Ahmad Fari Arief LopaNessuna valutazione finora
- Jurnal AcneDocumento5 pagineJurnal AcneStacy VaniaNessuna valutazione finora
- Design and Characterization of Flucytosine Loaded Bioadhesive in Situ Ophthalmic Gel For Improved BioavailabilityDocumento4 pagineDesign and Characterization of Flucytosine Loaded Bioadhesive in Situ Ophthalmic Gel For Improved BioavailabilityRam SahuNessuna valutazione finora
- Dandruff PDFDocumento8 pagineDandruff PDFArthur ChietraNessuna valutazione finora
- Chemical Peels in The Treatment of Acne PDFDocumento8 pagineChemical Peels in The Treatment of Acne PDFSpica AdharaNessuna valutazione finora
- Skin AntioksidanDocumento8 pagineSkin AntioksidanAngga SuryaNessuna valutazione finora
- Erythroderma (Generalized Exfoliative Dermatitis) - Background, Pathophysiology, EpidemiologyDocumento10 pagineErythroderma (Generalized Exfoliative Dermatitis) - Background, Pathophysiology, EpidemiologyAnonymous S0MyRHNessuna valutazione finora
- Leprosy Reaction: Oleh: Aryo Wibowo C11103224 Pembimbing: Dr. Cyntia Yulyana Supervisor: Dr. Muhlis, SP - KK, M.KesDocumento14 pagineLeprosy Reaction: Oleh: Aryo Wibowo C11103224 Pembimbing: Dr. Cyntia Yulyana Supervisor: Dr. Muhlis, SP - KK, M.Kesjoe ashikawaNessuna valutazione finora
- 07 Dermatological Formulation and Transdermal SystemsDocumento81 pagine07 Dermatological Formulation and Transdermal SystemsAlberto JacobusNessuna valutazione finora
- Antioxidant Activity and Phenolic Compounds in 32 Selected Herbs PDFDocumento10 pagineAntioxidant Activity and Phenolic Compounds in 32 Selected Herbs PDFMuhammad FaiqNessuna valutazione finora
- Basal Cell Carcinoma Diagnosis & TreatmentDocumento18 pagineBasal Cell Carcinoma Diagnosis & Treatmentjourey08Nessuna valutazione finora
- Eritroderma - Bolognia 4th EditionDocumento15 pagineEritroderma - Bolognia 4th EditionAisyahNessuna valutazione finora
- Bab I Pendahuluan 1.1 Latar Belakang: Case Report Session Ini Disusun Untuk Memenuhi Tugas Kepaniteraan KlinikDocumento33 pagineBab I Pendahuluan 1.1 Latar Belakang: Case Report Session Ini Disusun Untuk Memenuhi Tugas Kepaniteraan KlinikNurifnaNessuna valutazione finora
- Creams and Ointments, What's The Difference, Which Is BestDocumento3 pagineCreams and Ointments, What's The Difference, Which Is Bestchrizela20Nessuna valutazione finora
- Drug AllergyDocumento450 pagineDrug AllergyAlice CorsiNessuna valutazione finora
- Drug Allergy BookDocumento332 pagineDrug Allergy Bookmegah_asia13Nessuna valutazione finora
- 9 - Anti Fungal DrugsDocumento73 pagine9 - Anti Fungal Drugszynab123Nessuna valutazione finora
- Vesiculobullous DiseasesDocumento40 pagineVesiculobullous Diseasessgoeldoc_550661200100% (1)
- AHA in Treatment of Signs of AgingDocumento10 pagineAHA in Treatment of Signs of AgingChamomillaeFlosNessuna valutazione finora
- Different Types of Skin Lesions ExplainedDocumento7 pagineDifferent Types of Skin Lesions Explained_carido_Nessuna valutazione finora
- Scar KeliodDocumento38 pagineScar KeliodjoshuaNessuna valutazione finora
- Proton Pump InhibitorDocumento9 pagineProton Pump InhibitorAnkush BiswasNessuna valutazione finora
- Skin Diseases Affecting The VulvaDocumento7 pagineSkin Diseases Affecting The VulvaNoraNessuna valutazione finora
- Retinoids in TherapyDocumento52 pagineRetinoids in TherapyPanini PatankarNessuna valutazione finora
- Dermatomyositis: A Guide to Clinical Features, Diagnosis and ManagementDocumento26 pagineDermatomyositis: A Guide to Clinical Features, Diagnosis and Managementzaki100% (1)
- Guideline Konjungtivitis CPG-11 PDFDocumento33 pagineGuideline Konjungtivitis CPG-11 PDFhanumzniiNessuna valutazione finora
- Vesiculobullous Disorders in Children2Documento5 pagineVesiculobullous Disorders in Children2Luciana FernandesNessuna valutazione finora
- Lip Disorders: Cleft Lip and PalateDocumento59 pagineLip Disorders: Cleft Lip and Palatepasser by100% (1)
- Radiologis Gambaran Infeksi Tulang dan FrakturDocumento25 pagineRadiologis Gambaran Infeksi Tulang dan FrakturAnonymous HAbhRTs2TfNessuna valutazione finora
- Systemic Lupus Eritematus Kuliah KlerkDocumento76 pagineSystemic Lupus Eritematus Kuliah KlerkAnonymous y6NmP9NHkYNessuna valutazione finora
- Anti-Fungal Drugs Guide for Mycosis TreatmentDocumento6 pagineAnti-Fungal Drugs Guide for Mycosis Treatmentbilal ahmadNessuna valutazione finora
- Proton Pump Inhibitors v3Documento5 pagineProton Pump Inhibitors v3shwampaNessuna valutazione finora
- Spongiotic Dermatitis: Aka EczemaDocumento8 pagineSpongiotic Dermatitis: Aka EczemafadoNessuna valutazione finora
- Hordeulum, Chalazion, Pyogenic GranulomaDocumento4 pagineHordeulum, Chalazion, Pyogenic Granulomaaksy100% (1)
- Influence of Azelaic and Mandelic Acid Peels On SebumDocumento6 pagineInfluence of Azelaic and Mandelic Acid Peels On SebumRojas Evert AlonsoNessuna valutazione finora
- FLUID INTAKE AND HYDRATIONDocumento17 pagineFLUID INTAKE AND HYDRATIONVhiny ChaNessuna valutazione finora
- Kuliah Dermato Terapi 2013Documento46 pagineKuliah Dermato Terapi 2013Vania EssiandaNessuna valutazione finora
- Complications of Proton Pump Inhibitor Therapy 2017Documento14 pagineComplications of Proton Pump Inhibitor Therapy 2017Ingrid Miroslava Reyes DiazNessuna valutazione finora
- Antiplasmodial ActivityDocumento10 pagineAntiplasmodial ActivityAbhishek ShawNessuna valutazione finora
- Atopic Eczema 11Documento71 pagineAtopic Eczema 11khola aijazNessuna valutazione finora
- Chapter-03 Acneiform EruptionDocumento6 pagineChapter-03 Acneiform EruptionSylvia DiamondNessuna valutazione finora
- Tumor Jinak Ginekologi - 4Documento61 pagineTumor Jinak Ginekologi - 4Bayna LestariNessuna valutazione finora
- Bullous DiseaseDocumento42 pagineBullous DiseaseErika KusumawatiNessuna valutazione finora
- TX of StriaeDocumento13 pagineTX of StriaeadeNessuna valutazione finora
- RenovHyal Anti-Ageing Ingredient Reinforces Skin BarrierDocumento8 pagineRenovHyal Anti-Ageing Ingredient Reinforces Skin BarrierMaritaNetoNessuna valutazione finora
- Modul Tropis Mhs PDFDocumento12 pagineModul Tropis Mhs PDFAvabel Player 1Nessuna valutazione finora
- Hierarchical Archimedean Copulas Through Multivariate Compound DistributionsDocumento13 pagineHierarchical Archimedean Copulas Through Multivariate Compound DistributionsHaroon ImtiazNessuna valutazione finora
- Dislocation Nucleation Governed Softening and MaxiDocumento5 pagineDislocation Nucleation Governed Softening and MaxiHaroon ImtiazNessuna valutazione finora
- Master Thesis InterfaceDocumento71 pagineMaster Thesis InterfaceHaroon ImtiazNessuna valutazione finora
- LwmericalDocumento33 pagineLwmericalHaroon ImtiazNessuna valutazione finora
- A G I S: Dmission Uide FOR Nternational TudentsDocumento29 pagineA G I S: Dmission Uide FOR Nternational Tudentsমাহমুদ অর্ণবNessuna valutazione finora
- Theory Strength Anisotropic: General of ForDocumento23 pagineTheory Strength Anisotropic: General of ForHaroon ImtiazNessuna valutazione finora
- List of PublicationsDocumento1 paginaList of PublicationsHaroon ImtiazNessuna valutazione finora
- Net Positive Suction Head or NPSHDocumento4 pagineNet Positive Suction Head or NPSHsateesh chandNessuna valutazione finora
- Universities 19-4-2012Documento10 pagineUniversities 19-4-2012Haroon ImtiazNessuna valutazione finora
- Metrology - Wikipedia, The Free EncyclopediaDocumento11 pagineMetrology - Wikipedia, The Free EncyclopediaHaroon Imtiaz100% (1)
- Comparative Effect of Aqueous Aloe Vera Extracts On Oxidative Stress in Alloxan Induced Diabetic Wistar RatsDocumento6 pagineComparative Effect of Aqueous Aloe Vera Extracts On Oxidative Stress in Alloxan Induced Diabetic Wistar RatsEditor IJTSRDNessuna valutazione finora
- Washington State University nrf2 Study Raises Health Promoting Factors-NotesDocumento26 pagineWashington State University nrf2 Study Raises Health Promoting Factors-Notesapi-253513646100% (4)
- Effects of Postharvest Application of Methyl Jasmonate On Physicochemical Characteristics and Antioxidant System of The Blueberry FruitDocumento8 pagineEffects of Postharvest Application of Methyl Jasmonate On Physicochemical Characteristics and Antioxidant System of The Blueberry FruitFernando Herrero SinNessuna valutazione finora
- Raw Truth BookDocumento109 pagineRaw Truth Bookgungi1950100% (14)
- 6BBL0325 2 TheoriesofAgeingRandomDamageDocumento10 pagine6BBL0325 2 TheoriesofAgeingRandomDamageVaidehi KatariaNessuna valutazione finora
- Pitfalls, Problems, and Progress in Bronchopulmonary DysplasiaDocumento14 paginePitfalls, Problems, and Progress in Bronchopulmonary Dysplasiatrixie2k7Nessuna valutazione finora
- Fulic Acid Improve Maize Performance Water StressDocumento9 pagineFulic Acid Improve Maize Performance Water StressGerad ValdezNessuna valutazione finora
- Enzymatic and Non EnzymaticDocumento28 pagineEnzymatic and Non EnzymaticHaseen AhmedNessuna valutazione finora
- Radikal BebasDocumento20 pagineRadikal Bebasazzahra_hamidahNessuna valutazione finora
- Free Radicals: Understanding the BasicsDocumento4 pagineFree Radicals: Understanding the BasicsRavi PreethiNessuna valutazione finora
- Protandim peer-reviewed studies summaryDocumento1 paginaProtandim peer-reviewed studies summaryIe InterneXpertsNessuna valutazione finora
- Introduction to Cell Injury and Death in PathologyDocumento133 pagineIntroduction to Cell Injury and Death in PathologyMd. Zayed ZamanNessuna valutazione finora
- Asma Thesis Black Print 5 SetDocumento106 pagineAsma Thesis Black Print 5 SetHajveri Printing Services100% (1)
- Malina 2020Documento10 pagineMalina 2020Jaime VZNessuna valutazione finora
- Investigations of Riboflavin Photolysis Via Coloured Light in The Nitro Blue Tetrazolium Assay For Superoxide Dismutase ActivityDocumento8 pagineInvestigations of Riboflavin Photolysis Via Coloured Light in The Nitro Blue Tetrazolium Assay For Superoxide Dismutase ActivitySonny RawlsNessuna valutazione finora
- Molecules 24 04277 v2Documento28 pagineMolecules 24 04277 v2KosmetykiNessuna valutazione finora
- Paper 8699Documento13 paginePaper 8699IJARSCT JournalNessuna valutazione finora
- Effect of Sodium Metabisulfite On Lipid Peroxidation and Enzyme Activi-Ties in Adult Rat Stomach and SpleenDocumento8 pagineEffect of Sodium Metabisulfite On Lipid Peroxidation and Enzyme Activi-Ties in Adult Rat Stomach and Spleenام محمدNessuna valutazione finora
- Foliar Application of ZincDocumento6 pagineFoliar Application of ZincNelsonNessuna valutazione finora
- Hydrogen Peroxide TherapyDocumento31 pagineHydrogen Peroxide TherapySavoryHubbard100% (1)
- Cuscuta Chinensislam. - A Systematic Review On PDFDocumento17 pagineCuscuta Chinensislam. - A Systematic Review On PDFDintakurthi PrasanthNessuna valutazione finora
- Redox-Genome Interactions in Health and DiseaseDocumento650 pagineRedox-Genome Interactions in Health and Diseaseaurea_ramNessuna valutazione finora
- Nils Metzler-Nolte, Ulrich Schatzschneider - Bioinorganic Chemistry - A Practical Course-De Gruyter (2009)Documento149 pagineNils Metzler-Nolte, Ulrich Schatzschneider - Bioinorganic Chemistry - A Practical Course-De Gruyter (2009)shunmugamNessuna valutazione finora
- Role of Oxidative Stress On Chronic Kidney Disease ProgressionDocumento9 pagineRole of Oxidative Stress On Chronic Kidney Disease ProgressionMohammad WahyuNessuna valutazione finora
- Oxidative Stress and Antioxidants - Their Role in Human Disease-Nova Biomedical Books (2009)Documento374 pagineOxidative Stress and Antioxidants - Their Role in Human Disease-Nova Biomedical Books (2009)io montoya v100% (1)
- 2020 - Treatment of Melasma A Review of Less Commonly Used AntioxidantsDocumento8 pagine2020 - Treatment of Melasma A Review of Less Commonly Used Antioxidantsnancyerlen100% (1)
- Ameliorative Potential of Bambara Nuts (Vigna SubterraneDocumento12 pagineAmeliorative Potential of Bambara Nuts (Vigna SubterraneAustin AiraodionNessuna valutazione finora
- AminothiolsDocumento23 pagineAminothiolsKatsu MatasuNessuna valutazione finora
- Ageing Biology: The Causes of SenescenceDocumento45 pagineAgeing Biology: The Causes of SenescenceAbhilash Kumar MuthuramanNessuna valutazione finora
- I Am BarleyDocumento8 pagineI Am BarleyJohn Patrick MuzanyiNessuna valutazione finora