Potrebbero piacerti anche
- Meher Baba - Glimpses of The God-Man Part 6 (274p)Documento274 pagineMeher Baba - Glimpses of The God-Man Part 6 (274p)paraisoptptNessuna valutazione finora
- Meher Baba - Glimpses of The God-Man Part 2 (422p)Documento422 pagineMeher Baba - Glimpses of The God-Man Part 2 (422p)paraisoptptNessuna valutazione finora
- Six Systems - Max MullerDocumento524 pagineSix Systems - Max MullerparaisoptptNessuna valutazione finora
- The Paces of Yu, The Shaman's Steps and Ame-no-Ukihashi - The Art of NinzuwuDocumento6 pagineThe Paces of Yu, The Shaman's Steps and Ame-no-Ukihashi - The Art of NinzuwuparaisoptptNessuna valutazione finora
- The Origins of SufismDocumento6 pagineThe Origins of SufismparaisoptptNessuna valutazione finora
- Therapy 101207215845 Phpapp01Documento43 pagineTherapy 101207215845 Phpapp01paraisoptpt100% (1)
- Deepening Emotional Literacy, Robert A. Masters PDFDocumento125 pagineDeepening Emotional Literacy, Robert A. Masters PDFparaisoptpt100% (2)
- Diet Night ShadesDocumento45 pagineDiet Night ShadesparaisoptptNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Raw Vegan Food CombiningDocumento4 pagineRaw Vegan Food CombiningSimo100% (16)
- Freebie Bundle-50 Pages.Documento76 pagineFreebie Bundle-50 Pages.Kia MercadoNessuna valutazione finora
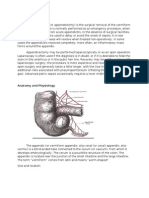
- AppendectomyDocumento5 pagineAppendectomyCris EstoniloNessuna valutazione finora
- Chemical Changes in Food During Processing and Storage: March 2016Documento3 pagineChemical Changes in Food During Processing and Storage: March 2016Pankti SutharNessuna valutazione finora
- Med Surge 3Documento55 pagineMed Surge 3rave andrei MacanayaNessuna valutazione finora
- 400+ Important Biology Questions and Answer PDF For SSC @WWW - Letsstudytogether.co PDFDocumento29 pagine400+ Important Biology Questions and Answer PDF For SSC @WWW - Letsstudytogether.co PDFShubham100% (1)
- Soal Explanation TextDocumento5 pagineSoal Explanation TextnadhifsaputroNessuna valutazione finora
- محاضرات الكيمياء كاملة PDFDocumento124 pagineمحاضرات الكيمياء كاملة PDFفراس الموسويNessuna valutazione finora
- THE DIGESTIVE SYSTEM 22 (Autosaved)Documento141 pagineTHE DIGESTIVE SYSTEM 22 (Autosaved)Jairus ChaloNessuna valutazione finora
- CHRONIC DIRRHEA FinalDocumento93 pagineCHRONIC DIRRHEA FinalAtifNessuna valutazione finora
- Ayurveda TreasuresDocumento54 pagineAyurveda TreasuresRajasimhan BaskarNessuna valutazione finora
- Apparent or True Digestibility: Protein Amino Acids Starch Glucose Fats Fatty Acids, MonoglyceridesDocumento13 pagineApparent or True Digestibility: Protein Amino Acids Starch Glucose Fats Fatty Acids, MonoglyceridesOyedotun TundeNessuna valutazione finora
- Maximum Marks: 40 Time Allotted: 50 Minutes Total Questions: 40 Choose The Correct Answer-Section A: Chemistry and BiologyDocumento2 pagineMaximum Marks: 40 Time Allotted: 50 Minutes Total Questions: 40 Choose The Correct Answer-Section A: Chemistry and Biologysapna sharmaNessuna valutazione finora
- Energy Sources For Fish and Calculating Enrgy DigestibilityDocumento26 pagineEnergy Sources For Fish and Calculating Enrgy DigestibilityVijay KarotiaNessuna valutazione finora
- Life ProcessesDocumento15 pagineLife ProcessesShalom LogosNessuna valutazione finora
- Lipid Digestion and AbsorptionDocumento72 pagineLipid Digestion and AbsorptionKavya GaneshNessuna valutazione finora
- Human Nutrition (Multiple Choice) QPDocumento17 pagineHuman Nutrition (Multiple Choice) QPkeenNessuna valutazione finora
- Amylase Production ThesisDocumento8 pagineAmylase Production Thesisafkodkedr100% (2)
- Pet Care of A Guinea Pig DifferentiatedDocumento16 paginePet Care of A Guinea Pig DifferentiatedandreiNessuna valutazione finora
- Gut Health in Poultry, 2013 PDFDocumento42 pagineGut Health in Poultry, 2013 PDFvetbcas100% (1)
- Second Periodic Test LatestDocumento20 pagineSecond Periodic Test LatestEimerej C. Spirit100% (1)
- Mammalian System-01 (Digestive System)Documento35 pagineMammalian System-01 (Digestive System)Saumya99Nessuna valutazione finora
- Soalan Science PMR Kertas 1 2006Documento21 pagineSoalan Science PMR Kertas 1 2006Azim SufieNessuna valutazione finora
- Allen: Pre-Medical: BiologyDocumento1 paginaAllen: Pre-Medical: BiologyDevdatta PatilNessuna valutazione finora
- IGCSE - Bio - Lesson Plan 4 - Food and DigestionDocumento4 pagineIGCSE - Bio - Lesson Plan 4 - Food and DigestionHisokagenNessuna valutazione finora
- Full Download Ebook Ebook PDF Our Social World Introduction To Sociology 7th Edition PDFDocumento41 pagineFull Download Ebook Ebook PDF Our Social World Introduction To Sociology 7th Edition PDFmichael.livengood88797% (39)
- Osmosis in The Digestive SystemDocumento1 paginaOsmosis in The Digestive SystemNadhiraAzhar_Nessuna valutazione finora
- NCM 103 Metab N EndocrineDocumento148 pagineNCM 103 Metab N EndocrinernrmmanphdNessuna valutazione finora
- Digestive System PowerPointDocumento35 pagineDigestive System PowerPointcelineNessuna valutazione finora
- Rabbit - WikipediaDocumento16 pagineRabbit - WikipediaAmy KlezanNessuna valutazione finora