Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Aaa 1Documento14 pagineAaa 1LunaPerfectNessuna valutazione finora
- Aaa 3Documento14 pagineAaa 3LunaPerfectNessuna valutazione finora
- Delivery CheckDocumento1 paginaDelivery CheckLunaPerfectNessuna valutazione finora
- The Natural History, Pathobiology, and Clinical Manifestations of Sars-Cov-2 InfectionsDocumento28 pagineThe Natural History, Pathobiology, and Clinical Manifestations of Sars-Cov-2 InfectionsJosé Luis Alonso EscamillaNessuna valutazione finora
- Asidosis Metabolik PDFDocumento6 pagineAsidosis Metabolik PDFafifurrahman_rizalNessuna valutazione finora
- Initial Assessment and ManagementDocumento21 pagineInitial Assessment and ManagementLunaPerfectNessuna valutazione finora
- MOSF PathogenesisDocumento8 pagineMOSF PathogenesisLunaPerfectNessuna valutazione finora
- Risk Factors For Type 2 Diabetes in Groups Stratified AccordingDocumento9 pagineRisk Factors For Type 2 Diabetes in Groups Stratified AccordingLunaPerfectNessuna valutazione finora
- CDC Growth Charts: United States: 95th 90th 75thDocumento1 paginaCDC Growth Charts: United States: 95th 90th 75thLunaPerfectNessuna valutazione finora
- Clinical Practice Guidelines For Antimicrobial Prophylaxis in Surgery (Update 2013)Documento89 pagineClinical Practice Guidelines For Antimicrobial Prophylaxis in Surgery (Update 2013)เด็กชายสมันตภัทร แฟนคลับอาจารย์กวงNessuna valutazione finora
- 2015 Trauma ManualDocumento165 pagine2015 Trauma ManualLunaPerfectNessuna valutazione finora
- Wound Healing ManualDocumento24 pagineWound Healing ManualaboujisNessuna valutazione finora
- Sugar-Sweetened Beverage Intake and Overweight in ChildrenDocumento7 pagineSugar-Sweetened Beverage Intake and Overweight in ChildrenLunaPerfectNessuna valutazione finora
- Kuisioner MinumanDocumento12 pagineKuisioner MinumanLunaPerfectNessuna valutazione finora
- AsriDocumento7 pagineAsriLunaPerfectNessuna valutazione finora
- Personal, Behavioral, and Environmental Risk and Protective Factors For Adolescent OverweightDocumento14 paginePersonal, Behavioral, and Environmental Risk and Protective Factors For Adolescent OverweightLunaPerfectNessuna valutazione finora
- CDC Growth Charts: United States: KG LB LBDocumento1 paginaCDC Growth Charts: United States: KG LB LBLunaPerfectNessuna valutazione finora
- CDC Growth Charts: United States: LB LB KGDocumento1 paginaCDC Growth Charts: United States: LB LB KGLunaPerfectNessuna valutazione finora
- Personal, Behavioral, and Environmental Risk and Protective Factors For Adolescent OverweightDocumento14 paginePersonal, Behavioral, and Environmental Risk and Protective Factors For Adolescent OverweightLunaPerfectNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Diabetes Transformation Summit 2022 Day 3Documento5 pagineDiabetes Transformation Summit 2022 Day 3Paul Ioan PopescuNessuna valutazione finora
- Quick Energy Boost Yoga PosesDocumento5 pagineQuick Energy Boost Yoga PosesTai CheeNessuna valutazione finora
- Depresia PostpartumDocumento18 pagineDepresia PostpartumIonela BogdanNessuna valutazione finora
- Reviewer Nutrimonth QuizbeeDocumento47 pagineReviewer Nutrimonth QuizbeeDale Lyko AbionNessuna valutazione finora
- Bath Uni Coursework Cover SheetDocumento8 pagineBath Uni Coursework Cover Sheetvtdvkkjbf100% (2)
- Manual Terapia DigitalDocumento38 pagineManual Terapia DigitalPaolaQuitian0% (1)
- Fundamentals of NursingDocumento50 pagineFundamentals of NursingGuruKPO67% (3)
- 10 12 15hematoo Edit 1Documento10 pagine10 12 15hematoo Edit 1Anonymous 18GsyXbNessuna valutazione finora
- Nicu ReportDocumento66 pagineNicu ReportEr Shah Rukh QadriNessuna valutazione finora
- Portal HypertensionDocumento25 paginePortal HypertensionJainab SiddiquiNessuna valutazione finora
- Perioperative Preparation and Management: Pre-OperativeDocumento14 paginePerioperative Preparation and Management: Pre-OperativeRed StohlNessuna valutazione finora
- Immunology MCQ BY: Dr. AlgassimDocumento4 pagineImmunology MCQ BY: Dr. AlgassimmohamedNessuna valutazione finora
- Classification of Trauma in ChildrenDocumento9 pagineClassification of Trauma in ChildrenRita ValenteNessuna valutazione finora
- Copd Action Plan PDFDocumento2 pagineCopd Action Plan PDFdtech2Nessuna valutazione finora
- Hex A Chloro But A DieneDocumento15 pagineHex A Chloro But A DieneAfraNessuna valutazione finora
- Language and ArtsDocumento17 pagineLanguage and ArtsOpium PoppyNessuna valutazione finora
- Hiperpigmentasi Paska InflamasiDocumento18 pagineHiperpigmentasi Paska InflamasiNoventyChristiNessuna valutazione finora
- FinalDocumento3 pagineFinalAnupam Saraf IINessuna valutazione finora
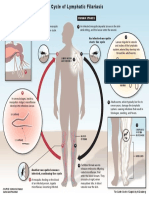
- Lymphatic Filariasis Life Cycle PDFDocumento1 paginaLymphatic Filariasis Life Cycle PDFlauraNessuna valutazione finora
- A Study To Evaluate The Effectiveness of Structured Teaching Program On Chemotherapy Administration in Terms of Knowledge Among Student Nurses in A Selected Institution at HubballiDocumento3 pagineA Study To Evaluate The Effectiveness of Structured Teaching Program On Chemotherapy Administration in Terms of Knowledge Among Student Nurses in A Selected Institution at HubballiInternational Journal of Innovative Science and Research TechnologyNessuna valutazione finora
- 9721 Smoking Cessaton Newsletter 1981Documento8 pagine9721 Smoking Cessaton Newsletter 1981Julia PurperaNessuna valutazione finora
- Training Manual PDFDocumento145 pagineTraining Manual PDFCalinnescuNessuna valutazione finora
- Hospital Environment For Sick ChildDocumento10 pagineHospital Environment For Sick ChildKhushbu KatariaNessuna valutazione finora
- Peds 2015-3501 Full PDFDocumento19 paginePeds 2015-3501 Full PDFJehan VahlepyNessuna valutazione finora
- Case Taking ProformaDocumento7 pagineCase Taking ProformaDr Dushyant Kamal Dhari50% (2)
- 1116005I Rev. 02Documento2 pagine1116005I Rev. 02kirubel demelashNessuna valutazione finora
- Target Product Profile Imbruvica - IbrutinibDocumento1 paginaTarget Product Profile Imbruvica - IbrutinibDiti ShahNessuna valutazione finora
- 2-NPSG Chapter OBS Jan2023Documento8 pagine2-NPSG Chapter OBS Jan2023Blanca LanderosNessuna valutazione finora
- Acute PancreatitisDocumento9 pagineAcute PancreatitisestefygomezsNessuna valutazione finora
- Pathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19) A ReviewDocumento13 paginePathophysiology, Transmission, Diagnosis, and Treatment of Coronavirus Disease 2019 (COVID-19) A ReviewBryan Diaz LopezNessuna valutazione finora