Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Method Statement FINALDocumento61 pagineMethod Statement FINALshareyhou67% (3)
- Study Notes - Google Project Management Professional CertificateDocumento4 pagineStudy Notes - Google Project Management Professional CertificateSWAPNIL100% (1)
- (Problem Books in Mathematics) Antonio Caminha Muniz Neto - An Excursion Through Elementary Mathematics, Volume III - Discrete Mathematics and Polynomial Algebra (2018, Springer)Documento647 pagine(Problem Books in Mathematics) Antonio Caminha Muniz Neto - An Excursion Through Elementary Mathematics, Volume III - Discrete Mathematics and Polynomial Algebra (2018, Springer)Anonymous iH6noeaX7100% (2)
- Assignment November11 KylaAccountingDocumento2 pagineAssignment November11 KylaAccountingADRIANO, Glecy C.Nessuna valutazione finora
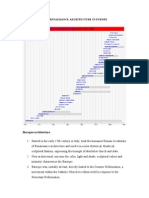
- Post Renaissance Architecture in EuropeDocumento10 paginePost Renaissance Architecture in Europekali_007Nessuna valutazione finora
- Protecting or Pilfering? Neoliberal Conservationist Marine Protected Areas in The Experience of Coastal Granada, The PhilippinesDocumento11 pagineProtecting or Pilfering? Neoliberal Conservationist Marine Protected Areas in The Experience of Coastal Granada, The PhilippinesWanderley JuniorNessuna valutazione finora
- Geoforum: George Holmes, Connor J. CavanaghDocumento11 pagineGeoforum: George Holmes, Connor J. CavanaghWanderley JuniorNessuna valutazione finora
- Neoliberalism in Ecotourism? The New Development Paradigm of Multinational Projects in AfricaDocumento15 pagineNeoliberalism in Ecotourism? The New Development Paradigm of Multinational Projects in AfricaWanderley JuniorNessuna valutazione finora
- Montes 2019Documento23 pagineMontes 2019Wanderley JuniorNessuna valutazione finora
- Deforestation and Reforestation of Latin America and The Caribbean (2001 - 2010)Documento10 pagineDeforestation and Reforestation of Latin America and The Caribbean (2001 - 2010)Wanderley JuniorNessuna valutazione finora
- Allaby 2006 - Tropical Rain Forests PDFDocumento289 pagineAllaby 2006 - Tropical Rain Forests PDFWanderley JuniorNessuna valutazione finora
- Microsoft Powerpoint BasicsDocumento20 pagineMicrosoft Powerpoint BasicsJonathan LocsinNessuna valutazione finora
- Assessing Apical PulseDocumento5 pagineAssessing Apical PulseMatthew Ryan100% (1)
- Department of Education: Raiseplus Weekly Plan For Blended LearningDocumento3 pagineDepartment of Education: Raiseplus Weekly Plan For Blended LearningMARILYN CONSIGNANessuna valutazione finora
- SCIENCE 11 WEEK 6c - Endogenic ProcessDocumento57 pagineSCIENCE 11 WEEK 6c - Endogenic ProcessChristine CayosaNessuna valutazione finora
- Worked Solution Paper5 A LevelDocumento8 pagineWorked Solution Paper5 A LevelBhoosan AncharazNessuna valutazione finora
- Monitor 14sepDocumento2 pagineMonitor 14sepabhaymvyas1144Nessuna valutazione finora
- Affidavit of Co OwnershipDocumento2 pagineAffidavit of Co OwnershipEmer MartinNessuna valutazione finora
- Ferobide Applications Brochure English v1 22Documento8 pagineFerobide Applications Brochure English v1 22Thiago FurtadoNessuna valutazione finora
- Driver Drowsiness Detection System Using Raspberry PiDocumento7 pagineDriver Drowsiness Detection System Using Raspberry PiIJRASETPublicationsNessuna valutazione finora
- CV Ashish PDFDocumento3 pagineCV Ashish PDFRoshan KejariwalNessuna valutazione finora
- Canon I-SENSYS MF411dw Parts CatalogDocumento79 pagineCanon I-SENSYS MF411dw Parts Catalogmarian100% (1)
- Current Harmonics: Electric Power System Power QualityDocumento3 pagineCurrent Harmonics: Electric Power System Power QualityAlliver SapitulaNessuna valutazione finora
- Tec066 6700 PDFDocumento2 pagineTec066 6700 PDFExclusivo VIPNessuna valutazione finora
- Subeeka Akbar Advance NutritionDocumento11 pagineSubeeka Akbar Advance NutritionSubeeka AkbarNessuna valutazione finora
- 2015 Student Handbook 16 January 2015Documento66 pagine2015 Student Handbook 16 January 2015John KhanNessuna valutazione finora
- What Is The PCB Shelf Life Extending The Life of PCBsDocumento9 pagineWhat Is The PCB Shelf Life Extending The Life of PCBsjackNessuna valutazione finora
- BAMDocumento111 pagineBAMnageswara_mutyalaNessuna valutazione finora
- Ethical Hacking IdDocumento24 pagineEthical Hacking IdSilvester Dian Handy PermanaNessuna valutazione finora
- Glossary of Blasting TermsDocumento13 pagineGlossary of Blasting TermsNitesh JainNessuna valutazione finora
- Febrile Neutropenia GuidelineDocumento8 pagineFebrile Neutropenia GuidelineAslesa Wangpathi PagehgiriNessuna valutazione finora
- Asus x453Documento5 pagineAsus x453Rhiry Ntuh AthryNessuna valutazione finora
- Fin Accounting IFRS 2e Ch13Documento62 pagineFin Accounting IFRS 2e Ch13Nguyễn Vinh QuangNessuna valutazione finora
- Dist - Propor.danfoss PVG32Documento136 pagineDist - Propor.danfoss PVG32Michal BujaraNessuna valutazione finora
- Agenda - 2 - Presentation - MS - IUT - Thesis Proposal PPT Muhaiminul 171051001Documento13 pagineAgenda - 2 - Presentation - MS - IUT - Thesis Proposal PPT Muhaiminul 171051001Tanvir AhmadNessuna valutazione finora
- DP-1520 PMDocumento152 pagineDP-1520 PMIon JardelNessuna valutazione finora