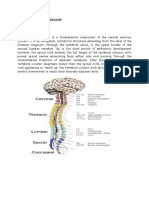
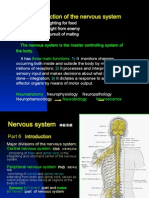
Potrebbero piacerti anche
- Macroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheDocumento14 pagineMacroscopic Neuroanatomy: Neuroanatomy Deals With The Structure of TheRoziel Anne BasaNessuna valutazione finora
- Excitable Cells: Monographs in Modern Biology for Upper School and University CoursesDa EverandExcitable Cells: Monographs in Modern Biology for Upper School and University CoursesNessuna valutazione finora
- Anatomy and Physiology The Spinal CordDocumento11 pagineAnatomy and Physiology The Spinal CordKatrina CaveNessuna valutazione finora
- Continuum Spinal Cord AnatomyDocumento25 pagineContinuum Spinal Cord AnatomyslojnotakNessuna valutazione finora
- Review of Anatomy and Physiology of CNS: Blood-Brain BarrierDocumento36 pagineReview of Anatomy and Physiology of CNS: Blood-Brain BarrierTimothy CaragNessuna valutazione finora
- 1 Hist Sept 13 ReadingNerve - Tissue2020Documento26 pagine1 Hist Sept 13 ReadingNerve - Tissue2020n-gorNessuna valutazione finora
- Thespinalcord: A Review of Functional NeuroanatomyDocumento18 pagineThespinalcord: A Review of Functional NeuroanatomyasfwegereNessuna valutazione finora
- Neuroanatomy, Cerebellum: Statpearls (Internet) - Treasure Island (FL) : Statpearls Publishing 2023 JanDocumento5 pagineNeuroanatomy, Cerebellum: Statpearls (Internet) - Treasure Island (FL) : Statpearls Publishing 2023 JanANISA RIFKA RIDHONessuna valutazione finora
- Nerve Tissue & Nervous System: Assoc. Prof Dr. Karim Al-Jashamy IMS/MSU 2010Documento27 pagineNerve Tissue & Nervous System: Assoc. Prof Dr. Karim Al-Jashamy IMS/MSU 2010Kaushik BhuvaNessuna valutazione finora
- Neuroanatomy Notes 2010Documento33 pagineNeuroanatomy Notes 2010Jennifer David InauenNessuna valutazione finora
- Rewiew FINAL 1Documento13 pagineRewiew FINAL 1achalbob18Nessuna valutazione finora
- Three Main Functions:: The Nervous System Is The Master Controlling System of The BodyDocumento34 pagineThree Main Functions:: The Nervous System Is The Master Controlling System of The Bodyapi-19641337Nessuna valutazione finora
- Chapter IIDocumento29 pagineChapter IIKurnia pralisaNessuna valutazione finora
- Acknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsDocumento7 pagineAcknowledgments: Gray's Anatomy, The Remarkable Work From Which Gray's Clinical Neuroanatomy IsAndreea LăzăroiuNessuna valutazione finora
- The Nervous System 2Documento84 pagineThe Nervous System 2Erika MatiasNessuna valutazione finora
- 02 - CNS DevelopmentDocumento3 pagine02 - CNS DevelopmentCemre KuzeyNessuna valutazione finora
- Neurology Base AlmostdocxDocumento76 pagineNeurology Base AlmostdocxAyad IdresNessuna valutazione finora
- Spinal NotesDocumento7 pagineSpinal NotesUsman Ali AkbarNessuna valutazione finora
- Chapter 3: Anatomy of The Spinal Cord: Skip To Main Contentskip To Navigation (Accesskey N)Documento12 pagineChapter 3: Anatomy of The Spinal Cord: Skip To Main Contentskip To Navigation (Accesskey N)SN WijesinheNessuna valutazione finora
- 09 Nerve TissueDocumento13 pagine09 Nerve TissueGovind V PNessuna valutazione finora
- The Structure of A Neuron J.kalatDocumento5 pagineThe Structure of A Neuron J.kalatSára VargaNessuna valutazione finora
- A Brief Introduction To Basic FunctionalDocumento50 pagineA Brief Introduction To Basic FunctionalkiflomNessuna valutazione finora
- 8 Nervous SystemDocumento7 pagine8 Nervous Systemangel.loking10Nessuna valutazione finora
- Table of Content: Seria L No Page NoDocumento48 pagineTable of Content: Seria L No Page NoHadia NoorNessuna valutazione finora
- Table of Content: Serial No Page NoDocumento33 pagineTable of Content: Serial No Page NoSID RAJPOOTNessuna valutazione finora
- Neuro Anatomy and Physiological Work BookDocumento36 pagineNeuro Anatomy and Physiological Work Bookshair e islamNessuna valutazione finora
- HLA7 - Nervous SyStemDocumento25 pagineHLA7 - Nervous SyStemMichael UrrutiaNessuna valutazione finora
- Rullida - EdullantesDocumento36 pagineRullida - EdullantesJaleah Gwyneth Fernandez EdullantesNessuna valutazione finora
- Bing MentahanDocumento15 pagineBing Mentahansecondfery2021Nessuna valutazione finora
- Centralnervoussystem CNSDocumento19 pagineCentralnervoussystem CNSÂhmęd ĐoolaNessuna valutazione finora
- Biology Lecture Notes: (Stemer'S Guide)Documento24 pagineBiology Lecture Notes: (Stemer'S Guide)Magdy Mohamed Abdallah AbdelkaderNessuna valutazione finora
- Lesson 3.Documento15 pagineLesson 3.Maedel Rose EsguerraNessuna valutazione finora
- Animal Nervous System: Logo HereDocumento32 pagineAnimal Nervous System: Logo HereAnne Claudette Capin TeofiloNessuna valutazione finora
- Brainstem and Nuclei Condensed Grayscale SlidesDocumento17 pagineBrainstem and Nuclei Condensed Grayscale SlidesMobarobberNessuna valutazione finora
- Nervous Tissue: Prof. Dr. Anas Sarwar QureshiDocumento12 pagineNervous Tissue: Prof. Dr. Anas Sarwar QureshiShafaqat Ghani Shafaqat GhaniNessuna valutazione finora
- Introduction and Spinal CordDocumento27 pagineIntroduction and Spinal Cordapi-196413370% (1)
- The Nervous SystemDocumento5 pagineThe Nervous SystemStephen YorNessuna valutazione finora
- Neuron and Glial CellsDocumento8 pagineNeuron and Glial CellsMurilo Bonet100% (1)
- Spinalcord and It's Connections Neuro Anatomy - 2Documento30 pagineSpinalcord and It's Connections Neuro Anatomy - 2Jamaru NishaNessuna valutazione finora
- Rullida - EdullantesDocumento36 pagineRullida - EdullantesJaleah Gwyneth Fernandez EdullantesNessuna valutazione finora
- L12 Nerves TissueDocumento8 pagineL12 Nerves Tissueعلي اياد شبيب حناشNessuna valutazione finora
- Psy 413 ProjectDocumento38 paginePsy 413 ProjectDumebi AneneNessuna valutazione finora
- Human BrainDocumento44 pagineHuman BrainSri vaishnava100% (1)
- Modern Review: Gross Anatomy of BrainDocumento32 pagineModern Review: Gross Anatomy of BrainVENU B ANessuna valutazione finora
- 2 NSDocumento65 pagine2 NSLilaNessuna valutazione finora
- Nervous TissueDocumento6 pagineNervous TissueAhmed JawdetNessuna valutazione finora
- Animal Physiology Organization of The Vertebrate Nervous System Animal BehaviorDocumento16 pagineAnimal Physiology Organization of The Vertebrate Nervous System Animal BehaviorAlyza MayoNessuna valutazione finora
- Brain Functions Underlying Speech - The Handbook of Phonetic Sciences - Blackwell Reference OnlineDocumento24 pagineBrain Functions Underlying Speech - The Handbook of Phonetic Sciences - Blackwell Reference OnlineHAKAN ÖZTEMÜRNessuna valutazione finora
- 1 - Spinal Cord NotesDocumento7 pagine1 - Spinal Cord NotesOceanChan QdNessuna valutazione finora
- Cerebellum TextDocumento7 pagineCerebellum TextJean Pierre Chastre LuzaNessuna valutazione finora
- CNS Histology 1Documento60 pagineCNS Histology 1naderipourkianaNessuna valutazione finora
- Nervous TissueDocumento14 pagineNervous Tissueyigermalamanuel32Nessuna valutazione finora
- Introduction and Organization of Nervous System: Presented by Dr. Amen FatimaDocumento37 pagineIntroduction and Organization of Nervous System: Presented by Dr. Amen FatimaShanailNessuna valutazione finora
- Nervous System HistologyDocumento4 pagineNervous System HistologyShelly Stephanie Bintoro100% (1)
- Oraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaDocumento10 pagineOraganisation of The Brain: Studenti (Gr. 13) : - Paun Cristina - Petrescu Andreea Elena - Poenaru Raluca SanzianaPaun CristinaNessuna valutazione finora
- NeuronDocumento23 pagineNeuronUnggul YudhaNessuna valutazione finora
- Trends in Neural EvolutionDocumento35 pagineTrends in Neural EvolutionTejpal Dahiya100% (1)
- Spinal Cord: StructureDocumento5 pagineSpinal Cord: StructureIka LestariNessuna valutazione finora
- Third Party AuditDocumento5 pagineThird Party AuditVijay IyerNessuna valutazione finora
- Debt Reduction - Large Indian Companies Are Now Obsessed With Debt ReductionDocumento5 pagineDebt Reduction - Large Indian Companies Are Now Obsessed With Debt ReductionVijay IyerNessuna valutazione finora
- Unit 1 Pfa Act and Rules: StructureDocumento101 pagineUnit 1 Pfa Act and Rules: StructureVijay IyerNessuna valutazione finora
- CET Zero To 100Documento1 paginaCET Zero To 100Vijay IyerNessuna valutazione finora
- Booklet On 8th Joint NoteDocumento40 pagineBooklet On 8th Joint NoteVijay IyerNessuna valutazione finora
- Regenration of Ventral Nerve Cord in Eisenia: FetidaDocumento6 pagineRegenration of Ventral Nerve Cord in Eisenia: FetidaVijay IyerNessuna valutazione finora
- Attachment Nmat Gurpreet Sir Lyst2719Documento5 pagineAttachment Nmat Gurpreet Sir Lyst2719Vijay IyerNessuna valutazione finora
- Cbroa News December 2020 2 PDFDocumento37 pagineCbroa News December 2020 2 PDFVijay IyerNessuna valutazione finora
- All India Bank Pensioners' & Retirees' Confederation (A.I.B.P.A.R.C.)Documento2 pagineAll India Bank Pensioners' & Retirees' Confederation (A.I.B.P.A.R.C.)Vijay IyerNessuna valutazione finora
- Advanced Control StrategiesDocumento2 pagineAdvanced Control StrategiesVijay IyerNessuna valutazione finora
- Interactions 2.1 Protein Ligand InteractionsDocumento6 pagineInteractions 2.1 Protein Ligand InteractionsVijay IyerNessuna valutazione finora
- 4.64 BE - Biotechnology Engg PDFDocumento42 pagine4.64 BE - Biotechnology Engg PDFVijay IyerNessuna valutazione finora
- Presentation of Learning in BacteriaDocumento21 paginePresentation of Learning in BacteriaVijay IyerNessuna valutazione finora
- Rapid Escape Response AssayDocumento1 paginaRapid Escape Response AssayVijay IyerNessuna valutazione finora
- 1 BioleachingDocumento424 pagine1 BioleachingAhmed AliNessuna valutazione finora
- Drug Desinging 4.1 Drug OptimizationDocumento8 pagineDrug Desinging 4.1 Drug OptimizationVijay IyerNessuna valutazione finora
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Da EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)Valutazione: 3 su 5 stelle3/5 (1)
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDa EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDValutazione: 5 su 5 stelle5/5 (4)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDa EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionValutazione: 4 su 5 stelle4/5 (404)
- The Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsDa EverandThe Ritual Effect: From Habit to Ritual, Harness the Surprising Power of Everyday ActionsValutazione: 4 su 5 stelle4/5 (4)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDa EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedValutazione: 4.5 su 5 stelle4.5/5 (82)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDa EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityValutazione: 4 su 5 stelle4/5 (35)
- Cult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryDa EverandCult, A Love Story: Ten Years Inside a Canadian Cult and the Subsequent Long Road of RecoveryValutazione: 4 su 5 stelle4/5 (46)
- The Obesity Code: Unlocking the Secrets of Weight LossDa EverandThe Obesity Code: Unlocking the Secrets of Weight LossValutazione: 4 su 5 stelle4/5 (6)
- By the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsDa EverandBy the Time You Read This: The Space between Cheslie's Smile and Mental Illness—Her Story in Her Own WordsNessuna valutazione finora
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDa EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeValutazione: 2 su 5 stelle2/5 (1)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsDa EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsValutazione: 4.5 su 5 stelle4.5/5 (170)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDa EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Raising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsDa EverandRaising Mentally Strong Kids: How to Combine the Power of Neuroscience with Love and Logic to Grow Confident, Kind, Responsible, and Resilient Children and Young AdultsValutazione: 5 su 5 stelle5/5 (1)
- The Twentysomething Treatment: A Revolutionary Remedy for an Uncertain AgeDa EverandThe Twentysomething Treatment: A Revolutionary Remedy for an Uncertain AgeValutazione: 4.5 su 5 stelle4.5/5 (2)
- Manipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesDa EverandManipulation: The Ultimate Guide To Influence People with Persuasion, Mind Control and NLP With Highly Effective Manipulation TechniquesValutazione: 4.5 su 5 stelle4.5/5 (1412)
- Summary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisDa EverandSummary: Outlive: The Science and Art of Longevity by Peter Attia MD, With Bill Gifford: Key Takeaways, Summary & AnalysisValutazione: 4.5 su 5 stelle4.5/5 (44)
- Summary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisDa EverandSummary: Limitless: Upgrade Your Brain, Learn Anything Faster, and Unlock Your Exceptional Life By Jim Kwik: Key Takeaways, Summary and AnalysisValutazione: 5 su 5 stelle5/5 (8)
- Dark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Da EverandDark Psychology & Manipulation: Discover How To Analyze People and Master Human Behaviour Using Emotional Influence Techniques, Body Language Secrets, Covert NLP, Speed Reading, and Hypnosis.Valutazione: 4.5 su 5 stelle4.5/5 (110)
- How to ADHD: The Ultimate Guide and Strategies for Productivity and Well-BeingDa EverandHow to ADHD: The Ultimate Guide and Strategies for Productivity and Well-BeingValutazione: 1 su 5 stelle1/5 (2)
- To Explain the World: The Discovery of Modern ScienceDa EverandTo Explain the World: The Discovery of Modern ScienceValutazione: 3.5 su 5 stelle3.5/5 (51)
- Self-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!Da EverandSelf-Care for Autistic People: 100+ Ways to Recharge, De-Stress, and Unmask!Valutazione: 5 su 5 stelle5/5 (1)
- The Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsDa EverandThe Garden Within: Where the War with Your Emotions Ends and Your Most Powerful Life BeginsNessuna valutazione finora
- Critical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsDa EverandCritical Thinking: How to Effectively Reason, Understand Irrationality, and Make Better DecisionsValutazione: 4.5 su 5 stelle4.5/5 (39)
- The Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeDa EverandThe Courage Habit: How to Accept Your Fears, Release the Past, and Live Your Courageous LifeValutazione: 4.5 su 5 stelle4.5/5 (254)