Potrebbero piacerti anche
- Mapa Conceptual Territorios de La Violencia (Conflictos Colombianos)Documento1 paginaMapa Conceptual Territorios de La Violencia (Conflictos Colombianos)Daniela Chilatra UrueñaNessuna valutazione finora
- Ciencias Económicas y PolíticasDocumento3 pagineCiencias Económicas y PolíticasDaniela Chilatra UrueñaNessuna valutazione finora
- Articulo Traducido Exposicion ParasitosDocumento7 pagineArticulo Traducido Exposicion ParasitosDaniela Chilatra UrueñaNessuna valutazione finora
- 5 Enzimas y Cinética Enzimática PDFDocumento12 pagine5 Enzimas y Cinética Enzimática PDFGabriela EscalanteNessuna valutazione finora
- Catedra de La PazDocumento2 pagineCatedra de La PazDaniela Chilatra UrueñaNessuna valutazione finora
- 4 - Capítulo I. Las Enzimas PDFDocumento99 pagine4 - Capítulo I. Las Enzimas PDFyulleyNessuna valutazione finora
- CARTILLA TUTORIAL INGLES 1 NuevaDocumento72 pagineCARTILLA TUTORIAL INGLES 1 NuevaDaniela Chilatra UrueñaNessuna valutazione finora
- Ciencias SocialesDocumento2 pagineCiencias SocialesDaniela Chilatra UrueñaNessuna valutazione finora
- Ciencias SocialesDocumento2 pagineCiencias SocialesDaniela Chilatra UrueñaNessuna valutazione finora
- Bioactive Peptides - En.esDocumento17 pagineBioactive Peptides - En.esDaniela Chilatra UrueñaNessuna valutazione finora
- Estudio Descriptivo de Los Danos Ambientales - DialnetDocumento21 pagineEstudio Descriptivo de Los Danos Ambientales - DialnetSandiivel CalderOnNessuna valutazione finora
- Articulo 16 de La ConstituciónDocumento1 paginaArticulo 16 de La ConstituciónDaniela Chilatra UrueñaNessuna valutazione finora
- La Cantante CalvaDocumento46 pagineLa Cantante CalvaQuiqueTar1Nessuna valutazione finora
- Expresión de EmocionesDocumento3 pagineExpresión de EmocionesDaniela Chilatra UrueñaNessuna valutazione finora
- 04 FernandezDocumento10 pagine04 FernandezDaniela Chilatra UrueñaNessuna valutazione finora
- CaracterizacionDeLaBiodiversidadAcuaticaYDeLaCalidad PDFDocumento7 pagineCaracterizacionDeLaBiodiversidadAcuaticaYDeLaCalidad PDFDaniela Chilatra UrueñaNessuna valutazione finora
- La Naturaleza Atomica de La MateriaDocumento19 pagineLa Naturaleza Atomica de La MateriaDaniela Chilatra UrueñaNessuna valutazione finora
- Franco - 1990 - Ecologia de Poblaciones PDFDocumento6 pagineFranco - 1990 - Ecologia de Poblaciones PDFDaniela Chilatra UrueñaNessuna valutazione finora
- Amazonia ColombianaDocumento2 pagineAmazonia ColombianaDaniela Chilatra UrueñaNessuna valutazione finora
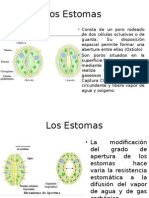
- EstomasDocumento11 pagineEstomasDaniela Chilatra UrueñaNessuna valutazione finora
- Manifestaciones de Afecto en Mi Comunidad EducativaDocumento4 pagineManifestaciones de Afecto en Mi Comunidad EducativaDaniela Chilatra Urueña86% (7)
- El Cuaderno 24 - 1Documento10 pagineEl Cuaderno 24 - 1Daniela Chilatra UrueñaNessuna valutazione finora
- Drosophila melanogaster mutantesDocumento2 pagineDrosophila melanogaster mutantesDaniela Chilatra Urueña100% (1)
- 8 - Amibas de Vida Libre en Seres Humanos - 2Documento13 pagine8 - Amibas de Vida Libre en Seres Humanos - 2Marco Antonio Ramirez VargasNessuna valutazione finora
- Diferencias Entre CélulasDocumento1 paginaDiferencias Entre CélulasDaniela Chilatra UrueñaNessuna valutazione finora
- Lep - Rhopalocera Clave de SuperfamiliasDocumento2 pagineLep - Rhopalocera Clave de SuperfamiliasDaniela Chilatra UrueñaNessuna valutazione finora
- Estructuras de Las BacteriasDocumento4 pagineEstructuras de Las BacteriasDaniela Chilatra UrueñaNessuna valutazione finora
- Diferencias Entre CélulasDocumento1 paginaDiferencias Entre CélulasDaniela Chilatra UrueñaNessuna valutazione finora
- 3.clasificacion de Las BacteriasDocumento39 pagine3.clasificacion de Las BacteriasMijail Soliz AlvarezNessuna valutazione finora
- Antología de Probabilidad y Estadística I PDFDocumento72 pagineAntología de Probabilidad y Estadística I PDFEsteban Mateus Williamson50% (2)
- Historia de La BellezaDocumento17 pagineHistoria de La BellezaRobotin CalderonNessuna valutazione finora
- Revalorando cultura peruanaDocumento4 pagineRevalorando cultura peruanaELEAZAR JESUS MOLINA ROJASNessuna valutazione finora
- Hds Latex PDFDocumento5 pagineHds Latex PDFVioleta Patricia Silva AstorgaNessuna valutazione finora
- Final Epidemiología LaboralDocumento10 pagineFinal Epidemiología LaboralVALMENNessuna valutazione finora
- Conclusiones de Diagnostico ComunitarioDocumento1 paginaConclusiones de Diagnostico ComunitarioRafael Villacorta50% (2)
- Fascículo 16. Panamá CCDocumento8 pagineFascículo 16. Panamá CCErwin OvNessuna valutazione finora
- Seguidor de Lineas Con 4 SensoresDocumento11 pagineSeguidor de Lineas Con 4 SensoresChuy RuizNessuna valutazione finora
- Emancipación y Organización Nacional. O. OszlakDocumento6 pagineEmancipación y Organización Nacional. O. OszlakLuciana MedinaNessuna valutazione finora
- Guía Investigación Adaptativa convocatoria 2 2016Documento7 pagineGuía Investigación Adaptativa convocatoria 2 2016Jose Luis LaraNessuna valutazione finora
- 7 y 8 EstadísticaDocumento6 pagine7 y 8 EstadísticaKarina Henostroza AyalaNessuna valutazione finora
- Formato de Flujo de EfectivoDocumento8 pagineFormato de Flujo de EfectivoSbeidy RoqueNessuna valutazione finora
- Plan Refuerzo Escolar 2024Documento16 paginePlan Refuerzo Escolar 2024percy83% (6)
- Disney o La Construcción DelDocumento11 pagineDisney o La Construcción DelJc MateusNessuna valutazione finora
- Congelacion de AlimentosDocumento9 pagineCongelacion de AlimentosEdson paitanNessuna valutazione finora
- Valoración Documental y Disposición Final de Los Documentos 2Documento15 pagineValoración Documental y Disposición Final de Los Documentos 2Jaime Andres Pinilla Torres45% (31)
- Resolucion de Direccion Ejecutiva-000377-2022-De - AnexosDocumento75 pagineResolucion de Direccion Ejecutiva-000377-2022-De - AnexosAndres RS ZKNessuna valutazione finora
- Concepto y Elementos Del MemorandumDocumento6 pagineConcepto y Elementos Del MemorandumAngelXtNessuna valutazione finora
- Regresión MúltipleDocumento34 pagineRegresión MúltipleLuigi ItaloNessuna valutazione finora
- Matriz EFE y EFIDocumento5 pagineMatriz EFE y EFIAnonymous bty2U5JryjNessuna valutazione finora
- Alerta Sanitaria Vacuna Vax-SpiralDocumento3 pagineAlerta Sanitaria Vacuna Vax-SpiralcarlosbaresNessuna valutazione finora
- Breviario de Estructura de Los MaterialesDocumento62 pagineBreviario de Estructura de Los MaterialesKatiaPeraltaNessuna valutazione finora
- SANITARIO ECOCLEAN SINGLE - FTC 008 1Documento2 pagineSANITARIO ECOCLEAN SINGLE - FTC 008 1Marjourie Denisse Urroz SacasaNessuna valutazione finora
- Guia-Escalas 2022-2Documento5 pagineGuia-Escalas 2022-2Mister AndersonNessuna valutazione finora
- Montecristi TesisDocumento58 pagineMontecristi TesisCristobal Argenis CruzNessuna valutazione finora
- ProblemarioDocumento17 pagineProblemarioQuique OrozcoNessuna valutazione finora
- Análisis comparado de tres planes estratégicos sectoriales en la Argentina post-convertibilidadDocumento187 pagineAnálisis comparado de tres planes estratégicos sectoriales en la Argentina post-convertibilidadCamelyn Katia Cadillo TiburcioNessuna valutazione finora
- Cómo Hacer Una Síntesis en Diseño GráficoDocumento7 pagineCómo Hacer Una Síntesis en Diseño GráficoRichard BastianNessuna valutazione finora
- CV Caceres Arpasi MalenaDocumento8 pagineCV Caceres Arpasi MalenaMalena Mariluna CaceresNessuna valutazione finora