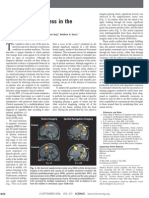
Potrebbero piacerti anche
- Malmo 1942Documento14 pagineMalmo 1942Víctor FuentesNessuna valutazione finora
- Perceptual Learning To Discriminate The Intensity and Spatial Location of Nociceptive Stimuli Mancini Et Al. 2016Documento10 paginePerceptual Learning To Discriminate The Intensity and Spatial Location of Nociceptive Stimuli Mancini Et Al. 2016somantammalwar99Nessuna valutazione finora
- Visuomotor Learning in Immersive 3D Virtual Reality in Parkinson's Disease and in AgingDocumento18 pagineVisuomotor Learning in Immersive 3D Virtual Reality in Parkinson's Disease and in AgingKristineNessuna valutazione finora
- Topographic Generalization of Tactile PeDocumento9 pagineTopographic Generalization of Tactile Pesomantammalwar99Nessuna valutazione finora
- 2007 Averbeck LeeDocumento8 pagine2007 Averbeck LeeLiya MaNessuna valutazione finora
- Garrigan (2008) Perceptual Learning Depends On Perceptual ConstancyDocumento6 pagineGarrigan (2008) Perceptual Learning Depends On Perceptual ConstancyJuan Francisco MuñozNessuna valutazione finora
- CrescentiniEtAl2011 PDFDocumento12 pagineCrescentiniEtAl2011 PDFdepisapiNessuna valutazione finora
- Lampiran 1 Permenpan2Documento13 pagineLampiran 1 Permenpan2RaodhatulJannahNessuna valutazione finora
- What Is NeurofeedbackDocumento30 pagineWhat Is NeurofeedbackgenerjustnNessuna valutazione finora
- Acute Exercise Improves Prefrontal Cortex But Not Hippocampal Function in Healthy AdultsDocumento11 pagineAcute Exercise Improves Prefrontal Cortex But Not Hippocampal Function in Healthy AdultsAlexander ElkoninNessuna valutazione finora
- The Role of Attention in Visual Processing: John H. R. Maunsell and Erik P. CookDocumento10 pagineThe Role of Attention in Visual Processing: John H. R. Maunsell and Erik P. CookrifaripotNessuna valutazione finora
- SCIENCE 2006 Detecting Awareness in The Vegetative StateDocumento1 paginaSCIENCE 2006 Detecting Awareness in The Vegetative StatepreghieraperlavitaNessuna valutazione finora
- Rapid Learning of A Phonemic Discrimination in The First Hours of LifeDocumento13 pagineRapid Learning of A Phonemic Discrimination in The First Hours of LifeTammysNessuna valutazione finora
- Ifa PercepcionDocumento12 pagineIfa PercepcionNathalie OrtizNessuna valutazione finora
- DelanoRobles 2007Documento8 pagineDelanoRobles 2007Carmen DélanoNessuna valutazione finora
- Deprivation-Induced Plasticity in The Early Central Circuits of The Rodent Visual, Auditory, and Olfactory Systems 2024Documento21 pagineDeprivation-Induced Plasticity in The Early Central Circuits of The Rodent Visual, Auditory, and Olfactory Systems 2024anacarolinadepaduaaNessuna valutazione finora
- Basso Etal++2015Documento11 pagineBasso Etal++2015Violeta Aguayo MüllerNessuna valutazione finora
- (Art) MRI SCANNER Symbolic Representations in Motor Sequence LearningDocumento10 pagine(Art) MRI SCANNER Symbolic Representations in Motor Sequence LearningPriss SaezNessuna valutazione finora
- Dinstein Autism Neuron 2010Documento9 pagineDinstein Autism Neuron 2010blocheNessuna valutazione finora
- Wildes Et Al-2014-International Journal of Eating DisordersDocumento4 pagineWildes Et Al-2014-International Journal of Eating DisordersThodorisMaurogiannidisNessuna valutazione finora
- The Differing Impact of Multisensory and Unisensory Integration On BehaviorDocumento6 pagineThe Differing Impact of Multisensory and Unisensory Integration On BehaviorD.PassmoreNessuna valutazione finora
- The Neural Correlates of Conscious VisionDocumento14 pagineThe Neural Correlates of Conscious VisionKarla ValderramaNessuna valutazione finora
- Biological MotionDocumento14 pagineBiological MotionchysaNessuna valutazione finora
- Steinemann-Etal 2016 VisualSequenceP300Documento31 pagineSteinemann-Etal 2016 VisualSequenceP300Monica Arias HigueraNessuna valutazione finora
- Clinical Neurophysiology: Kiwako Sakamoto, Hiroki Nakata, Ryusuke KakigiDocumento10 pagineClinical Neurophysiology: Kiwako Sakamoto, Hiroki Nakata, Ryusuke KakigiValeria Alejandra Hilda Riquelme CárcamoNessuna valutazione finora
- Yarmohammadi-Samani Et AlDocumento18 pagineYarmohammadi-Samani Et AlFrancisco MoraNessuna valutazione finora
- Endurance Exercise Selectively Impairs Prefrontal-Dependent CognitionDocumento9 pagineEndurance Exercise Selectively Impairs Prefrontal-Dependent CognitionhowardNessuna valutazione finora
- Bimodal (Auditory and Visual) Left Frontoparietal Circuitry For Sensorimotor Integration and Sensorimotor LearningDocumento9 pagineBimodal (Auditory and Visual) Left Frontoparietal Circuitry For Sensorimotor Integration and Sensorimotor LearningcokkyNessuna valutazione finora
- Perceptual Learning and Adult Cortical Plasticity2009Journal of PhysiologyDocumento9 paginePerceptual Learning and Adult Cortical Plasticity2009Journal of Physiologyshefaligupta91Nessuna valutazione finora
- Kanske 2010Documento10 pagineKanske 2010童帅Nessuna valutazione finora
- Society For Research in Child Development, Wiley Child DevelopmentDocumento25 pagineSociety For Research in Child Development, Wiley Child Developmentcecilia martinezNessuna valutazione finora
- Bola Proceedingsofthe National Academyof Sciences 2017Documento16 pagineBola Proceedingsofthe National Academyof Sciences 2017courursulaNessuna valutazione finora
- Loss of Consciousness Is Associated With Stabilization of Cortical ActivityDocumento12 pagineLoss of Consciousness Is Associated With Stabilization of Cortical ActivityFelly LiuNessuna valutazione finora
- Ruysschaert 2013Documento6 pagineRuysschaert 2013NéstorNessuna valutazione finora
- Walker Nature 2003Documento5 pagineWalker Nature 2003canoneinversoNessuna valutazione finora
- Environmental Enrichment and NeurogenesisDocumento7 pagineEnvironmental Enrichment and NeurogenesisAdriana CravoNessuna valutazione finora
- 1 s2.0 S0028393221001354 MainDocumento10 pagine1 s2.0 S0028393221001354 MainSimone MaucciNessuna valutazione finora
- 3251 FullDocumento10 pagine3251 FullChaoran ChengNessuna valutazione finora
- Overview CerebellumDocumento21 pagineOverview CerebellumwilsonnianNessuna valutazione finora
- Chen CB2019Documento12 pagineChen CB2019Juan ChenNessuna valutazione finora
- Case Study: Unreliable Evoked Responses in AutismDocumento11 pagineCase Study: Unreliable Evoked Responses in AutismEdward Ventura BarrientosNessuna valutazione finora
- Functional Cerebral Reorganization Following Motor Sequence Learning Through Mental Practice With Motor ImageryDocumento10 pagineFunctional Cerebral Reorganization Following Motor Sequence Learning Through Mental Practice With Motor ImageryMayrita Antonio UrquidiNessuna valutazione finora
- The Critical Role of Sleep Spindles in Hippocampal-Dependent Memory: A Pharmacology StudyDocumento11 pagineThe Critical Role of Sleep Spindles in Hippocampal-Dependent Memory: A Pharmacology StudySharan SahotaNessuna valutazione finora
- Studying The Dynamics of Autonomic Activity During Emotional ExperienceDocumento11 pagineStudying The Dynamics of Autonomic Activity During Emotional Experiencepsic.tatianebezerraNessuna valutazione finora
- Boosting Human Learning by HypnosisDocumento5 pagineBoosting Human Learning by HypnosisIsabelNessuna valutazione finora
- Mecanism of AttentionDocumento21 pagineMecanism of Attention'Minerva AlejandraNessuna valutazione finora
- Basso Etal.2015 PDFDocumento12 pagineBasso Etal.2015 PDFSixto Guardo MarmolNessuna valutazione finora
- A Balancing Act: Physical Balance, Through Arousal, in Uences Size PerceptionDocumento28 pagineA Balancing Act: Physical Balance, Through Arousal, in Uences Size Perceptionعلي عباسNessuna valutazione finora
- Validity of The Postrotary Nystagmus Test For Measuring Vestibular FunctionDocumento8 pagineValidity of The Postrotary Nystagmus Test For Measuring Vestibular FunctionTasneem BismillaNessuna valutazione finora
- Ambiguous Figures - What Happens in The Brain When Perception Changes But Not The StimulusDocumento23 pagineAmbiguous Figures - What Happens in The Brain When Perception Changes But Not The StimulusPriyanka ChakrabortyNessuna valutazione finora
- 1963 Mountcastle-Muscle Spindles? PDFDocumento28 pagine1963 Mountcastle-Muscle Spindles? PDFSilvio SánchezNessuna valutazione finora
- Review: The Reorienting System of The Human Brain: From Environment To Theory of MindDocumento19 pagineReview: The Reorienting System of The Human Brain: From Environment To Theory of MindMarina Sánchez PicazoNessuna valutazione finora
- Electrophysiological Precursors of Social ConformityDocumento8 pagineElectrophysiological Precursors of Social Conformitypsic.tatianebezerraNessuna valutazione finora
- Learning Brain RythmDocumento13 pagineLearning Brain RythmDa CuNessuna valutazione finora
- Rokem 2009Documento6 pagineRokem 2009feopazoNessuna valutazione finora
- Effects of Chewing On Cognitive Processing Speed PDFDocumento6 pagineEffects of Chewing On Cognitive Processing Speed PDFValeria Alejandra Hilda Riquelme CárcamoNessuna valutazione finora
- Dound Indice Flashs Illutsion SDTDocumento8 pagineDound Indice Flashs Illutsion SDTangieNessuna valutazione finora
- György Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)Documento20 pagineGyörgy Buzsáki - The Brain From Inside Out-Oxford University Press, USA (2019)juannnnNessuna valutazione finora
- Intelligence: Daniela Nussbaumer, Roland H. Grabner, Elsbeth SternDocumento13 pagineIntelligence: Daniela Nussbaumer, Roland H. Grabner, Elsbeth SternZim ShahNessuna valutazione finora
- HBO2 For FlapsDocumento8 pagineHBO2 For Flapstonylee24Nessuna valutazione finora
- BAby It's Cold Outside (Duet)Documento4 pagineBAby It's Cold Outside (Duet)tonylee24Nessuna valutazione finora
- 2005 J Physiol-Brubakk-Single Air Dive Endothelial FunctionDocumento6 pagine2005 J Physiol-Brubakk-Single Air Dive Endothelial FunctionTony LeeNessuna valutazione finora
- Troponin T Vs Troponin I ComparisonDocumento9 pagineTroponin T Vs Troponin I Comparisontonylee1962Nessuna valutazione finora
- IBS-Q For HCPDocumento2 pagineIBS-Q For HCPtonylee24Nessuna valutazione finora
- Jingle Bell Rock - SATBDocumento6 pagineJingle Bell Rock - SATBtonylee24100% (1)
- Musical Experience Shapes Human Brainstem Encoding of Linguistic Pitch PatternsDocumento7 pagineMusical Experience Shapes Human Brainstem Encoding of Linguistic Pitch Patternstonylee24Nessuna valutazione finora
- Smoking Cessation in RSNDocumento5 pagineSmoking Cessation in RSNtonylee24Nessuna valutazione finora
- Hyperbaric Oxygen For Post Concussive SyndromeDocumento28 pagineHyperbaric Oxygen For Post Concussive Syndrometonylee24Nessuna valutazione finora
- The Mechanism of Speech Processing in Congenital Amusia: Evidence From Mandarin SpeakersDocumento11 pagineThe Mechanism of Speech Processing in Congenital Amusia: Evidence From Mandarin Speakerstonylee24Nessuna valutazione finora
- The Mechanism of Speech Processing in Congenital Amusia: Evidence From Mandarin SpeakersDocumento11 pagineThe Mechanism of Speech Processing in Congenital Amusia: Evidence From Mandarin Speakerstonylee24Nessuna valutazione finora
- Widespread Auditory Deficits in Tune DeafnessDocumento29 pagineWidespread Auditory Deficits in Tune Deafnesstonylee24Nessuna valutazione finora
- Hyperbaric Oxygen For Post Concussive SyndromeDocumento28 pagineHyperbaric Oxygen For Post Concussive Syndrometonylee24Nessuna valutazione finora
- Effects of Culture On Musical Pitch PerceptionDocumento8 pagineEffects of Culture On Musical Pitch Perceptiontonylee24Nessuna valutazione finora
- Relating Pitch Awareness To Phonemic Awareness in Children: Implications For Tone-Deafness and DyslexiaDocumento5 pagineRelating Pitch Awareness To Phonemic Awareness in Children: Implications For Tone-Deafness and Dyslexiatonylee24Nessuna valutazione finora
- A Dual-Stream Neuroanatomy of SingingDocumento15 pagineA Dual-Stream Neuroanatomy of Singingtonylee24Nessuna valutazione finora
- Musical Experience Shapes Human Brainstem Encoding of Linguistic Pitch PatternsDocumento7 pagineMusical Experience Shapes Human Brainstem Encoding of Linguistic Pitch Patternstonylee24Nessuna valutazione finora
- Pitch Perception and Production in Congenital Amusia: Evidence From Cantonese SpeakersDocumento13 paginePitch Perception and Production in Congenital Amusia: Evidence From Cantonese Speakerstonylee24Nessuna valutazione finora
- Phonological Processing in Adults With Deficits in Musical Pitch RecognitionDocumento13 paginePhonological Processing in Adults With Deficits in Musical Pitch Recognitiontonylee24Nessuna valutazione finora
- Tone-Deafness - A New Disconnection Syndrome?Documento14 pagineTone-Deafness - A New Disconnection Syndrome?tonylee24Nessuna valutazione finora
- Congenital Amusia (Or Tone-Deafness) Interferes With Pitch Processing in Tone LanguagesDocumento15 pagineCongenital Amusia (Or Tone-Deafness) Interferes With Pitch Processing in Tone Languagestonylee24Nessuna valutazione finora
- Tone Language Fluency Impairs Pitch DiscriminationDocumento5 pagineTone Language Fluency Impairs Pitch Discriminationtonylee24Nessuna valutazione finora
- Brainstem Encoding of Speech and Musical StimDocumento19 pagineBrainstem Encoding of Speech and Musical Stimtonylee24Nessuna valutazione finora
- Cross-Language Perception of Non-Native Tonal Contrasts: Effects of Native Phonological and Phonetic InfluencesDocumento20 pagineCross-Language Perception of Non-Native Tonal Contrasts: Effects of Native Phonological and Phonetic Influencestonylee24Nessuna valutazione finora
- The Linked Dual Representation Model of Vocal Perception and ProductionDocumento12 pagineThe Linked Dual Representation Model of Vocal Perception and Productiontonylee24Nessuna valutazione finora
- Action-Perception Mismatch in Tone-DeafnessDocumento5 pagineAction-Perception Mismatch in Tone-Deafnesstonylee24Nessuna valutazione finora
- Am J Hum Gen 2000 PDFDocumento4 pagineAm J Hum Gen 2000 PDFtonylee24Nessuna valutazione finora
- Musicians and Tone-Language Speakers Share Enhanced Brainstem Encoding But Not Perceptual Benefits For Musical PitchDocumento23 pagineMusicians and Tone-Language Speakers Share Enhanced Brainstem Encoding But Not Perceptual Benefits For Musical Pitchtonylee24Nessuna valutazione finora
- The Genetics of Congenital Amusia (Tone Deafness) : A Family-Aggregation StudyDocumento7 pagineThe Genetics of Congenital Amusia (Tone Deafness) : A Family-Aggregation Studytonylee24Nessuna valutazione finora
- Cog Quizzes 1 - 12Documento6 pagineCog Quizzes 1 - 12Frank TaiNessuna valutazione finora
- 131A LECTURES - ch0 5Documento257 pagine131A LECTURES - ch0 5Fatimah ZananaNessuna valutazione finora
- Neuro OphthalmologyDocumento702 pagineNeuro OphthalmologyFauziah HafshohNessuna valutazione finora
- On The Role of Spatial Phase and Phase Correlation in Vision, Illusion, and CognitionDocumento14 pagineOn The Role of Spatial Phase and Phase Correlation in Vision, Illusion, and Cognitiondw wwNessuna valutazione finora
- Theoretical Understanding of Early Visual ProcessesDocumento33 pagineTheoretical Understanding of Early Visual ProcessesVignesh A KNessuna valutazione finora
- TNN2008 Luo ChaominpublishedDocumento21 pagineTNN2008 Luo ChaominpublishedAsd fASFAS SNessuna valutazione finora
- Chapter 3Documento102 pagineChapter 3Womens Program Assosciation100% (1)
- 1general SensationDocumento141 pagine1general SensationAzimuddeen AzeliNessuna valutazione finora
- Review: Conscious Processing and The Global Neuronal Workspace HypothesisDocumento23 pagineReview: Conscious Processing and The Global Neuronal Workspace HypothesisSebastian Gonzalez MazoNessuna valutazione finora
- Distributed RepresentationsDocumento35 pagineDistributed RepresentationsabhinavNessuna valutazione finora
- Biological MotionDocumento14 pagineBiological MotionchysaNessuna valutazione finora
- Neuronal Dynamics From Single Neurons To Networks and Models of CognitionDocumento76 pagineNeuronal Dynamics From Single Neurons To Networks and Models of CognitionOrchidaceae PhalaenopsisNessuna valutazione finora
- Wagor 1980 (Retinotopic Organization of Striate and Extrastriate Visual Cortex in The Mouse) PDFDocumento16 pagineWagor 1980 (Retinotopic Organization of Striate and Extrastriate Visual Cortex in The Mouse) PDFFRANCISCO ELI LEZAMA GUTIERREZNessuna valutazione finora
- Binocular VisionDocumento35 pagineBinocular Visiongargi.c100% (3)
- Principles of Nerual ScienceDocumento28 paginePrinciples of Nerual ScienceRaunak MukherjeeNessuna valutazione finora
- Li Zhaoping-Understanding Vision - Theory, Models, and Data-Oxford University Press (2014)Documento397 pagineLi Zhaoping-Understanding Vision - Theory, Models, and Data-Oxford University Press (2014)paul mNessuna valutazione finora
- Welcome To CSE/NEUBEH 528: Computational Neuroscience: Today's AgendaDocumento27 pagineWelcome To CSE/NEUBEH 528: Computational Neuroscience: Today's AgendaEdward Kofi AzeleorNessuna valutazione finora
- Psychobiology Ch. 6: Vision Lecture NotesDocumento23 paginePsychobiology Ch. 6: Vision Lecture NotesGabriel CardozaNessuna valutazione finora
- Engineering Applications of Artificial Intelligence: Saijie Fan, Wei Liang, Derui Ding, Hui YuDocumento11 pagineEngineering Applications of Artificial Intelligence: Saijie Fan, Wei Liang, Derui Ding, Hui YuSameer AhmedNessuna valutazione finora
- Physiological Mechanisms of Color Vision Israel AbramovDocumento29 paginePhysiological Mechanisms of Color Vision Israel AbramovinvencaodemorelNessuna valutazione finora
- The Physiology of Human Vision: Steven M. LavalleDocumento17 pagineThe Physiology of Human Vision: Steven M. LavalleAdson AlcantaraNessuna valutazione finora
- RetinaDocumento12 pagineRetinagdudex118811Nessuna valutazione finora
- Receptive FieldsDocumento121 pagineReceptive FieldsGeorge BenghiaNessuna valutazione finora
- 6.Radar-Vision Fusion For Object ClassificationDocumento7 pagine6.Radar-Vision Fusion For Object ClassificationThanh LeNessuna valutazione finora
- Paper 1Documento14 paginePaper 1royalatlas.stashNessuna valutazione finora
- Perception Ch2-3 - The Eye and RetinaDocumento70 paginePerception Ch2-3 - The Eye and Retina楊楨琳Nessuna valutazione finora
- Bio Psychology Lecture Notes Chapter 6 The Visual SystemDocumento7 pagineBio Psychology Lecture Notes Chapter 6 The Visual SystemGeneric_Persona50% (2)
- A Simple Baseline For Video Restoration With Grouped Spatial-Temporal ShiftDocumento14 pagineA Simple Baseline For Video Restoration With Grouped Spatial-Temporal Shift黃丰楷Nessuna valutazione finora
- The EyeDocumento36 pagineThe EyeMuresan Radu CatalinNessuna valutazione finora
- 02 Higherlevel Visual ProcessingDocumento48 pagine02 Higherlevel Visual ProcessingsethNessuna valutazione finora