Potrebbero piacerti anche
- PM Risk For DummiesDocumento52 paginePM Risk For DummiesFelix Rinaldi100% (8)
- 009 Installation of Pumps Risk AssessmentDocumento2 pagine009 Installation of Pumps Risk Assessmentgangadharan000100% (13)
- Do You Have Esp FinalDocumento20 pagineDo You Have Esp FinalVrasidas Poulopoulos50% (2)
- (Herbert Kitschelt, Steven I. Wilkinson) Patrons, PDFDocumento387 pagine(Herbert Kitschelt, Steven I. Wilkinson) Patrons, PDFClemente Tomás Sánchez BarjaNessuna valutazione finora
- Screening and Identification of Laccase Producing Fungi From Environmental SamplesDocumento8 pagineScreening and Identification of Laccase Producing Fungi From Environmental SamplesUMYU Journal of Microbiology Research (UJMR)Nessuna valutazione finora
- Screening Isolation and Characterization of Cellulase Producing Micro-Organisms From SoilDocumento7 pagineScreening Isolation and Characterization of Cellulase Producing Micro-Organisms From SoilinventionjournalsNessuna valutazione finora
- A Comparative GC-MS Analysis of Bioactive Secondary Metabolites Produced by HalotolerantDocumento16 pagineA Comparative GC-MS Analysis of Bioactive Secondary Metabolites Produced by HalotolerantAmineJaouedNessuna valutazione finora
- 13 - 15v5i2 - 4 Serratia Marcescens OU50TDocumento5 pagine13 - 15v5i2 - 4 Serratia Marcescens OU50TIsworo RukmiNessuna valutazione finora
- Antimicrobial Potentials of Phospholipid Compound Produced by Halophilic Bacillus SubtilisDocumento6 pagineAntimicrobial Potentials of Phospholipid Compound Produced by Halophilic Bacillus Subtiliskerkour-abd1523Nessuna valutazione finora
- Agunbiade Et Al.. 2019. Evaluation of Fresh Water Actinomycete Bioflocculant and Its Biotechnological Applications in Wastewaters TreatmDocumento15 pagineAgunbiade Et Al.. 2019. Evaluation of Fresh Water Actinomycete Bioflocculant and Its Biotechnological Applications in Wastewaters TreatmAzb 711Nessuna valutazione finora
- Elkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondDocumento9 pagineElkady Et Al.. 2011. Bacillus Mojavensis Strain 32A, A Bioflocculant-Producing Bacterium Isolated From An Egyptian Salt Production PondAzb 711Nessuna valutazione finora
- Systematic Characterization of Potential Cellulolytic Marine Actinobacteria Actinoalloteichus SP Mha15 PDFDocumento7 pagineSystematic Characterization of Potential Cellulolytic Marine Actinobacteria Actinoalloteichus SP Mha15 PDFIT InventoryNessuna valutazione finora
- Bajaj, Singhal. 2011. Flocculation Properties of Poly (γ-Glutamic Acid) Produced from Bacillus subtilis IsolateDocumento8 pagineBajaj, Singhal. 2011. Flocculation Properties of Poly (γ-Glutamic Acid) Produced from Bacillus subtilis IsolateAzb 711Nessuna valutazione finora
- Degradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusDocumento7 pagineDegradation Kraft Indulin Lignin by Viridosporus Streptomyces BadiusMuhammad UmerNessuna valutazione finora
- Heliyon: Marwan Jawad Msarah, Izyanti Ibrahim, Aidil Abdul Hamid, Wan Syaidatul AqmaDocumento9 pagineHeliyon: Marwan Jawad Msarah, Izyanti Ibrahim, Aidil Abdul Hamid, Wan Syaidatul AqmaOlusegun OlasugbaNessuna valutazione finora
- Giri Et Al.. 2015. Production and Characterization of A Thermostable Bioflocculant From Bacillus Subtilis F9, Isolated From Wastewater SludgeDocumento6 pagineGiri Et Al.. 2015. Production and Characterization of A Thermostable Bioflocculant From Bacillus Subtilis F9, Isolated From Wastewater SludgeAzb 711Nessuna valutazione finora
- 603 1259 1 PBCVCXVDocumento6 pagine603 1259 1 PBCVCXVMaria AspriNessuna valutazione finora
- BIOTECh Research Paper 2022Documento28 pagineBIOTECh Research Paper 2022Duge Galtsa100% (1)
- Contrera Et Al 2013 NMRDocumento10 pagineContrera Et Al 2013 NMRSurajit BhattacharjeeNessuna valutazione finora
- Fungal Pretreatment of Agricultural Residues For Bioethanol ProductionDocumento7 pagineFungal Pretreatment of Agricultural Residues For Bioethanol ProductionDiegoVasqzNessuna valutazione finora
- Optimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaDocumento8 pagineOptimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaUMYU Journal of Microbiology Research (UJMR)Nessuna valutazione finora
- Pilot Scale Bioconversion of Rice and SuDocumento9 paginePilot Scale Bioconversion of Rice and SuPrassu SwtyNessuna valutazione finora
- Reuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuraDocumento9 pagineReuteri 2-20B and Pediococcus Acidilactici 0-11A Isolated From FuramostecocNessuna valutazione finora
- 7 - 36 - 42 - Omotosho Et Al., - 24Documento6 pagine7 - 36 - 42 - Omotosho Et Al., - 24RezanovianingrumNessuna valutazione finora
- Research Article: Isolation of Cellulose-Degrading Bacteria and Determination of Their Cellulolytic PotentialDocumento5 pagineResearch Article: Isolation of Cellulose-Degrading Bacteria and Determination of Their Cellulolytic PotentialSubs KatsNessuna valutazione finora
- Bacteriocin Production by Lactic Acid Bacteria Encapsulated in Calcium Alginate BeadsDocumento8 pagineBacteriocin Production by Lactic Acid Bacteria Encapsulated in Calcium Alginate BeadsangelicaaragonNessuna valutazione finora
- Protease Production by An Actinomycete MA1-1Documento7 pagineProtease Production by An Actinomycete MA1-1salin MajumdarNessuna valutazione finora
- Biotechnology & Biotechnological Equipment: A A B ADocumento5 pagineBiotechnology & Biotechnological Equipment: A A B AgrandisrfNessuna valutazione finora
- BBRC Vol 14 No 04 2021-20Documento7 pagineBBRC Vol 14 No 04 2021-20Dr Sharique AliNessuna valutazione finora
- Produccion de Laccasa Por Pleurotus OstreatusDocumento9 pagineProduccion de Laccasa Por Pleurotus OstreatusGABRIEL DE JESUS MONTES FUENTESNessuna valutazione finora
- Phylogenetically Diverse Acetaldehyde-Degrading Bacterial Community Deep Sea Water West Pacific OceanDocumento11 paginePhylogenetically Diverse Acetaldehyde-Degrading Bacterial Community Deep Sea Water West Pacific OceanAndrea EscobarNessuna valutazione finora
- DownloadDocumento8 pagineDownloadMizra Nurqalby NurdinNessuna valutazione finora
- Vaishali Randive, Et AlDocumento8 pagineVaishali Randive, Et AlOENDRIL DASNessuna valutazione finora
- Liu2019 Article Co-cultureOfBacillusCoagulansADocumento6 pagineLiu2019 Article Co-cultureOfBacillusCoagulansAGUIDO ERNESTO VILLOTA CALVACHINessuna valutazione finora
- Pan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.Documento5 paginePan, X., Chen, F., Wu, T., Tang, H., Dan Zhao, Z. 2009. The Acid, Bile Tolerance and Antimicrobial Property of Lactobacillus Acidophilus NIT. J. Food Control. 20 598-602.AfdhalRuslanNessuna valutazione finora
- Continuous Volatile Fatty Acid Production From Lignocellulosic Biomass by A Novel Rumen-Mimetic BioprocessDocumento6 pagineContinuous Volatile Fatty Acid Production From Lignocellulosic Biomass by A Novel Rumen-Mimetic BioprocessGuillermo IdarragaNessuna valutazione finora
- Enhanced Production of Lutein in Heterotrophic: Chlorella Protothecoides by Oxidative StressDocumento6 pagineEnhanced Production of Lutein in Heterotrophic: Chlorella Protothecoides by Oxidative StressPaola MarticorenaNessuna valutazione finora
- 1 Ijbrdfeb20171Documento8 pagine1 Ijbrdfeb20171TJPRC PublicationsNessuna valutazione finora
- Biotechnology ReportsDocumento6 pagineBiotechnology ReportsKevin PaterninaNessuna valutazione finora
- 23 15 15 DrinkingwaterDocumento7 pagine23 15 15 DrinkingwaterPrecious Marie AgabeNessuna valutazione finora
- 23 15 15 DrinkingwaterDocumento7 pagine23 15 15 DrinkingwaterPrecious Marie AgabeNessuna valutazione finora
- Purification and Characterization of A Bacteriocin Produced by Lactobacillus Lactis Isolated From Marine EnvironmentDocumento7 paginePurification and Characterization of A Bacteriocin Produced by Lactobacillus Lactis Isolated From Marine EnvironmentHoàng-NguyênĐặngNessuna valutazione finora
- Optimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaDocumento8 pagineOptimization of Culture Condition in The Production of Bioenzymes by Bacteria Isolated From Poultry Waste in Sokoto State, NigeriaUMYU Journal of Microbiology Research (UJMR)Nessuna valutazione finora
- Biochemistry ArticleDocumento7 pagineBiochemistry ArticleUli AsariNessuna valutazione finora
- A Novel Thiol-Dependent Serine Protease From Neocosmospora Sp. N1Documento12 pagineA Novel Thiol-Dependent Serine Protease From Neocosmospora Sp. N1Fatema MatkawalaNessuna valutazione finora
- Microbiological Research: Yanbing Zhu, Liyun Wu, Yanhong Chen, Hui Ni, Anfeng Xiao, Huinong CaiDocumento10 pagineMicrobiological Research: Yanbing Zhu, Liyun Wu, Yanhong Chen, Hui Ni, Anfeng Xiao, Huinong CaiJosé Larragaña OsunaNessuna valutazione finora
- HTTP WWW - Sciencedirect.com Science Ob MImg& Imagekey B6T6P-4VJ0CWR-5-7& Cdi 5036& User 687350& Search& CoverDate 06/30/2009& Ie SdarticleDocumento5 pagineHTTP WWW - Sciencedirect.com Science Ob MImg& Imagekey B6T6P-4VJ0CWR-5-7& Cdi 5036& User 687350& Search& CoverDate 06/30/2009& Ie SdarticlePaola CastroNessuna valutazione finora
- Lihong Zhao - Wenli Yin - Laccase Producing Strains - Protoplast FusionDocumento4 pagineLihong Zhao - Wenli Yin - Laccase Producing Strains - Protoplast FusionmllabateNessuna valutazione finora
- Immobilization of Rhodococcus Rhodochrous BX2 (An AcetonitriledegradingDocumento7 pagineImmobilization of Rhodococcus Rhodochrous BX2 (An AcetonitriledegradingSahar IrankhahNessuna valutazione finora
- Research Article: Effect of Cry Protein Induced Toxicity On Different Biochemical Parameters of The Tissue of InterestDocumento5 pagineResearch Article: Effect of Cry Protein Induced Toxicity On Different Biochemical Parameters of The Tissue of InterestArpita Khamrai KhanNessuna valutazione finora
- Biocontrol Potential of Lipopeptide Bio Surfactant Produced by Bacillus Sp. From Mangrove Ecosystem at Vellar-Coleroon Estuarine Complex Against Early Blight of TomatoDocumento9 pagineBiocontrol Potential of Lipopeptide Bio Surfactant Produced by Bacillus Sp. From Mangrove Ecosystem at Vellar-Coleroon Estuarine Complex Against Early Blight of TomatoInternational Journal of Innovative Science and Research TechnologyNessuna valutazione finora
- 6 PB PDFDocumento12 pagine6 PB PDFAkmalia Solehah 2203135247Nessuna valutazione finora
- Jurnal Teknologi: I - A - I A E B A M SDocumento5 pagineJurnal Teknologi: I - A - I A E B A M SrahayuNessuna valutazione finora
- Short Communication Characterization of Bioflocculant Producing-Bacteria Isolated From Tapioca Waste WaterDocumento4 pagineShort Communication Characterization of Bioflocculant Producing-Bacteria Isolated From Tapioca Waste WaterPrisca AyuNessuna valutazione finora
- Vishnuvardhan Reddy 2008 Isolation of PHA Bacteria From SewagesludgeDocumento7 pagineVishnuvardhan Reddy 2008 Isolation of PHA Bacteria From SewagesludgeASWINI J SNessuna valutazione finora
- Isolation and Characterization of Extracellular Thermoalkaline Protease Producing Bacillus Cereus Isolated From Tannery EffluentDocumento7 pagineIsolation and Characterization of Extracellular Thermoalkaline Protease Producing Bacillus Cereus Isolated From Tannery EffluenttheijesNessuna valutazione finora
- Varsha Shivaji Kulkarni and Bhagyashree Shashikant KulkarniDocumento11 pagineVarsha Shivaji Kulkarni and Bhagyashree Shashikant KulkarniCyriel Jade MendelloreNessuna valutazione finora
- Research Article Bacillus SubtilisDocumento10 pagineResearch Article Bacillus SubtilisDương Nguyễn Thùy DungNessuna valutazione finora
- Gong Et Al.. 2008. Bioflocculant Production by Culture of Serratia Ficaria and Its Application in Wastewater TreatmentDocumento7 pagineGong Et Al.. 2008. Bioflocculant Production by Culture of Serratia Ficaria and Its Application in Wastewater TreatmentAzb 711Nessuna valutazione finora
- Evaluation of The Antimicrobial Potential of Actinobacteria Strains Isolated From Mangrove Soils in The Municipality of São Caetano de Odivelas - Pará, BrazilDocumento5 pagineEvaluation of The Antimicrobial Potential of Actinobacteria Strains Isolated From Mangrove Soils in The Municipality of São Caetano de Odivelas - Pará, BrazilMamta AgarwalNessuna valutazione finora
- Acid Other MusDocumento9 pagineAcid Other MusRoberto CastellanosNessuna valutazione finora
- 61 12 653 PDFDocumento10 pagine61 12 653 PDFDavien UtoyoNessuna valutazione finora
- Isolation and Identification of Antimicrobial-Producing Lactic Acid Bacteria From Fermented CucumberDocumento9 pagineIsolation and Identification of Antimicrobial-Producing Lactic Acid Bacteria From Fermented CucumberAndrea MarceloNessuna valutazione finora
- crmb725 732 PDFDocumento9 paginecrmb725 732 PDFrababNessuna valutazione finora
- Study The Effects of Ethidium Bromide, Sds and Elevated Temperature On Stability of Multiple Antibiotic Resistances Plasmids ofDocumento15 pagineStudy The Effects of Ethidium Bromide, Sds and Elevated Temperature On Stability of Multiple Antibiotic Resistances Plasmids ofrababNessuna valutazione finora
- Partial Purification and Characterization of Protease IV From Pseudomonas AeruginosaDocumento13 paginePartial Purification and Characterization of Protease IV From Pseudomonas AeruginosarababNessuna valutazione finora
- Isolation and Identification of Antibiotic-Producing Actinom YcetesDocumento8 pagineIsolation and Identification of Antibiotic-Producing Actinom YcetesrababNessuna valutazione finora
- A Tribute To Fernando L. L. B. Carneiro (1913 - 2001)Documento2 pagineA Tribute To Fernando L. L. B. Carneiro (1913 - 2001)Roberto Urrutia100% (1)
- Unit 9 Caring and Sharing Year 2 KSSR Lesson PlanDocumento6 pagineUnit 9 Caring and Sharing Year 2 KSSR Lesson PlanopheliavioletNessuna valutazione finora
- JAIM InstructionsDocumento11 pagineJAIM InstructionsAlphaRaj Mekapogu100% (1)
- The Seven Perceptual Learning StylesDocumento3 pagineThe Seven Perceptual Learning StylesRamona FloreaNessuna valutazione finora
- Waves Scattering Induced by An Interface Crack in A Coated MaterialDocumento6 pagineWaves Scattering Induced by An Interface Crack in A Coated MaterialDarioLampaNessuna valutazione finora
- Purposive Comm. Group 6 PPT Concise Vers.Documento77 paginePurposive Comm. Group 6 PPT Concise Vers.rovicrosales1Nessuna valutazione finora
- 27 EtdsDocumento29 pagine27 EtdsSuhag PatelNessuna valutazione finora
- Tcps 2 Final WebDocumento218 pagineTcps 2 Final WebBornaGhannadiNessuna valutazione finora
- Practice Makes Perfect English Conversation UNIT 9Documento10 paginePractice Makes Perfect English Conversation UNIT 9Thaís C C MirandaNessuna valutazione finora
- Kolban's Book On C.H.I.P.Documento317 pagineKolban's Book On C.H.I.P.thiagocabral88Nessuna valutazione finora
- Healthcare ProfessionalismDocumento28 pagineHealthcare ProfessionalismAnjo CincoNessuna valutazione finora
- CBSE Class 10 Science Sample Paper-08 (For 2013)Documento11 pagineCBSE Class 10 Science Sample Paper-08 (For 2013)cbsesamplepaperNessuna valutazione finora
- Specification of SGP InterlayerDocumento3 pagineSpecification of SGP InterlayerHAN HANNessuna valutazione finora
- Information Sharing QuestionnaireDocumento7 pagineInformation Sharing Questionnairesheelasahoo0% (1)
- CHEM Test 2 SolutionsDocumento5 pagineCHEM Test 2 Solutionscuongtran_siegenNessuna valutazione finora
- 02.03.2017 SR Mechanical Engineer Externe Vacature InnoluxDocumento1 pagina02.03.2017 SR Mechanical Engineer Externe Vacature InnoluxMichel van WordragenNessuna valutazione finora
- TASK 500 Technical Memorandum No. 503 Review and Assessment of Drainage Control Policies, Procedures, AND Guidelines Final DraftDocumento31 pagineTASK 500 Technical Memorandum No. 503 Review and Assessment of Drainage Control Policies, Procedures, AND Guidelines Final DraftGreg BraswellNessuna valutazione finora
- Brandi Jones-5e-Lesson-Plan 2Documento3 pagineBrandi Jones-5e-Lesson-Plan 2api-491136095Nessuna valutazione finora
- Ieee Sight 2018 Call For PapersDocumento4 pagineIeee Sight 2018 Call For PapersThushar TomNessuna valutazione finora
- Herzlich Willkommen - Welcome: German Information Session For New ParentsDocumento20 pagineHerzlich Willkommen - Welcome: German Information Session For New ParentssammlissNessuna valutazione finora
- Organizational Behavior - Motivational Theories at Mcdonald's ReportDocumento11 pagineOrganizational Behavior - Motivational Theories at Mcdonald's ReportvnbioNessuna valutazione finora
- Non-Display Industrial Computers: User ManualDocumento100 pagineNon-Display Industrial Computers: User ManualJason100% (1)
- Bab 4 Ba501 Pecahan SeparaDocumento24 pagineBab 4 Ba501 Pecahan SeparaPrince ShanNessuna valutazione finora
- HowToCreate DataDriven HorizonCube OpendTectv4.4 PDFDocumento49 pagineHowToCreate DataDriven HorizonCube OpendTectv4.4 PDFKrisna Hanjar PrastawaNessuna valutazione finora
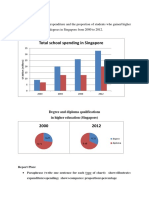
- Total School Spending in Singapore: Task 1 020219Documento6 pagineTotal School Spending in Singapore: Task 1 020219viper_4554Nessuna valutazione finora
- Listening Test 1.1Documento2 pagineListening Test 1.1momobear152Nessuna valutazione finora