Potrebbero piacerti anche
- KP-210217-Recruitment Consultant - FINAL RevDocumento1 paginaKP-210217-Recruitment Consultant - FINAL RevMaulitaNessuna valutazione finora
- Spatial Uncertainty of A Geoid Undulation Model in Guayaquil, EcuadorDocumento11 pagineSpatial Uncertainty of A Geoid Undulation Model in Guayaquil, EcuadorMaulitaNessuna valutazione finora
- 1 GOCE IntroductionDocumento62 pagine1 GOCE IntroductionMaulitaNessuna valutazione finora
- Alternating Current Circuits and Electromagnetic WavesDocumento65 pagineAlternating Current Circuits and Electromagnetic WavesMaulitaNessuna valutazione finora
- Spatial Uncertainty of A Geoid Undulation Model in Guayaquil, EcuadorDocumento11 pagineSpatial Uncertainty of A Geoid Undulation Model in Guayaquil, EcuadorMaulitaNessuna valutazione finora
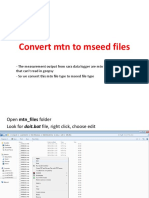
- Convert mtn files to mseed for dispersion analysisDocumento46 pagineConvert mtn files to mseed for dispersion analysisMaulitaNessuna valutazione finora
- Claprood Etal BSSA 2011Documento22 pagineClaprood Etal BSSA 2011Jennifer WatsonNessuna valutazione finora
- MMK Bab 5Documento96 pagineMMK Bab 5MaulitaNessuna valutazione finora
- Naskah Soal UN Fisika SMA 2012 Paket A81Documento13 pagineNaskah Soal UN Fisika SMA 2012 Paket A81agungfiristNessuna valutazione finora
- Long-Term Influence of Feeding Barley Treated With Lactic Acid and Heat On Performance and Energy Balance in Dairy CowsDocumento14 pagineLong-Term Influence of Feeding Barley Treated With Lactic Acid and Heat On Performance and Energy Balance in Dairy CowsMaulitaNessuna valutazione finora
- Effects of Dietary Addition of Cellulase and A Saccharomyces Cerevisiae Fermentation Product On Nutrient Digestibility, Rumen Fermentation and Enteric Methane Emissions in Growing GoatsDocumento16 pagineEffects of Dietary Addition of Cellulase and A Saccharomyces Cerevisiae Fermentation Product On Nutrient Digestibility, Rumen Fermentation and Enteric Methane Emissions in Growing GoatsMaulitaNessuna valutazione finora
- Effects of The Replacement of Concentrate and Fibre-Rich Hay by High-Quality Hay On Chewing, Rumination and Nutrient Digestibility in Non-Lactating Holstein CowsDocumento17 pagineEffects of The Replacement of Concentrate and Fibre-Rich Hay by High-Quality Hay On Chewing, Rumination and Nutrient Digestibility in Non-Lactating Holstein CowsMaulitaNessuna valutazione finora
- Chtest8 SolutionDocumento9 pagineChtest8 SolutionMaulitaNessuna valutazione finora
- Tabel ParabolaDocumento2 pagineTabel ParabolaMaulitaNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Alternative Feeds GuideDocumento128 pagineAlternative Feeds GuideKadiwala Dairy Farm100% (1)
- AnnotatedDocumento9 pagineAnnotatedapi-483714607Nessuna valutazione finora
- Spelling List 4Documento70 pagineSpelling List 4BobbingtonionNessuna valutazione finora
- Alternative Plant Protein Sources For Pigs and ChickensDocumento23 pagineAlternative Plant Protein Sources For Pigs and ChickensalvinsernaNessuna valutazione finora
- SARA em RuminantesDocumento12 pagineSARA em RuminantesGustavo SchneidersNessuna valutazione finora
- Practical guide to dairy cattle nutrition and feeding managementDocumento74 paginePractical guide to dairy cattle nutrition and feeding managementUmesh AdigaNessuna valutazione finora
- Ps The Complete Book of BariaurDocumento133 paginePs The Complete Book of Bariaurpoisonink100% (2)
- M. M. Rahman - A Review of Oxalate Poisoning in Domestic Animals Tolerance and Performance AspectsDocumento10 pagineM. M. Rahman - A Review of Oxalate Poisoning in Domestic Animals Tolerance and Performance AspectsNayara YoNessuna valutazione finora
- 3rd Term s1 Agricultural Science 1Documento41 pagine3rd Term s1 Agricultural Science 1Adelowo DanielNessuna valutazione finora
- Biology BrochureDocumento2 pagineBiology Brochureapi-317164998Nessuna valutazione finora
- Biodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusDocumento7 pagineBiodegradation of Agricultural Wastes (Rice Straw and Sorghum Stalk) Into Substrates of Utilizable Products Using White Rot FungusInes HrastinskiNessuna valutazione finora
- Online English, Maths and Science Revision TestsDocumento8 pagineOnline English, Maths and Science Revision Testsshailesh mahtoNessuna valutazione finora
- ThesisDocumento64 pagineThesisHamdi ShaarNessuna valutazione finora
- Nutrient Requirements For CattleDocumento50 pagineNutrient Requirements For CattleMohaajanan AliNessuna valutazione finora
- Important animal feedstuffs and their classificationDocumento20 pagineImportant animal feedstuffs and their classificationFAZALNessuna valutazione finora
- Rumen MicrofloraDocumento14 pagineRumen MicrofloraDrRameem BlochNessuna valutazione finora
- Teeth 131109091818 Phpapp01Documento42 pagineTeeth 131109091818 Phpapp01ariyatiNessuna valutazione finora
- Voluntary Food Intake and Diet Selection in Farm Animals - J.M. Forbes - 2nd Ed - 2007 - (CABI)Documento462 pagineVoluntary Food Intake and Diet Selection in Farm Animals - J.M. Forbes - 2nd Ed - 2007 - (CABI)José Pedro Casagrande TrentínNessuna valutazione finora
- Anatomy and Physiology of The Goat PDFDocumento7 pagineAnatomy and Physiology of The Goat PDFHannan MirzaNessuna valutazione finora
- Methane Mitigation Strategies From Livestock by SK Asraf HossainDocumento78 pagineMethane Mitigation Strategies From Livestock by SK Asraf HossainKandice Burgess100% (1)
- Effect of Dietary Buffers on Milk Production and Physiology in Early LactationDocumento8 pagineEffect of Dietary Buffers on Milk Production and Physiology in Early LactationNICOL GIORJINA LOPEZ HUILLCANessuna valutazione finora
- Ch 2 MCQ Questions Answers Digestive System Human Body Organs Functions ProcessesDocumento24 pagineCh 2 MCQ Questions Answers Digestive System Human Body Organs Functions ProcessesRamesh KulkarniNessuna valutazione finora
- SSS 1 ANIMAL HUSBANDRY EXAM REVIEWDocumento2 pagineSSS 1 ANIMAL HUSBANDRY EXAM REVIEWejogheneta100% (3)
- Digestive PowerpointDocumento44 pagineDigestive PowerpointMauricio BeltranNessuna valutazione finora
- Feed Additives in Ruminant Nutrition FINALDocumento39 pagineFeed Additives in Ruminant Nutrition FINALjss_bustamanteNessuna valutazione finora
- How Sustainable Is Organic Agriculture in The Phils Rodel G. Maghirang Et AlDocumento180 pagineHow Sustainable Is Organic Agriculture in The Phils Rodel G. Maghirang Et AlAbner Obniala Venus Jr.Nessuna valutazione finora
- Fsa 3038 PDFDocumento6 pagineFsa 3038 PDFGenna Prama NugrohoNessuna valutazione finora
- Probiotic Use in Sheep FeedDocumento5 pagineProbiotic Use in Sheep FeedRiska AulyaNessuna valutazione finora
- Leafy Corn Silage Info GuideDocumento24 pagineLeafy Corn Silage Info GuideMargo Lee100% (1)
- Grade 11 TestDocumento5 pagineGrade 11 TestClinton O'NeilNessuna valutazione finora