Potrebbero piacerti anche
- Ian D. Young (Author) - Introduction To Risk Calculation in Genetic Counseling, Third Edition-Oxford University Press, USA (2006) PDFDocumento252 pagineIan D. Young (Author) - Introduction To Risk Calculation in Genetic Counseling, Third Edition-Oxford University Press, USA (2006) PDFOmar AskanderNessuna valutazione finora
- Sarah Baker - The Alexander TechniqueDocumento125 pagineSarah Baker - The Alexander TechniquezameershahNessuna valutazione finora
- BiochemistryDocumento113 pagineBiochemistryMohammed Faizuddin siddiqui100% (1)
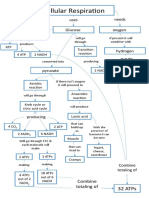
- Cellular Respiration Concept MapDocumento1 paginaCellular Respiration Concept MapmawakizakiNessuna valutazione finora
- Biochemistry Laboratory Modern Theory and Techniques-2nd EdDocumento78 pagineBiochemistry Laboratory Modern Theory and Techniques-2nd EdCharles EdwardNessuna valutazione finora
- Introduction To BiochemistryDocumento122 pagineIntroduction To Biochemistryqwerty123100% (1)
- ElectrolytesDocumento76 pagineElectrolytesanjilinbraganzaNessuna valutazione finora
- Interesting Q&A in Immunology - Part IIIDocumento21 pagineInteresting Q&A in Immunology - Part IIIVytheeshwaran Vedagiri100% (14)
- Test Bank For The Dynamic Child 1st Canadian Edition Franklin R Manis Alissa PencerDocumento22 pagineTest Bank For The Dynamic Child 1st Canadian Edition Franklin R Manis Alissa Pencerleahjamescwkqpobnet100% (40)
- A History of Membrane Transport and Bioenergetics PDFDocumento383 pagineA History of Membrane Transport and Bioenergetics PDFEdmilson RodriguesNessuna valutazione finora
- Enzymes LectureDocumento115 pagineEnzymes LectureJane Tai100% (1)
- EnzymesDocumento19 pagineEnzymesIS99057Nessuna valutazione finora
- EnzymesDocumento10 pagineEnzymesSanchit Dhyani100% (1)
- EnzymesDocumento20 pagineEnzymesDr.P.Natarajan100% (1)
- Biochemistry IntroductionDocumento54 pagineBiochemistry IntroductionNia Coline Macala MendozaNessuna valutazione finora
- What Are Enzymes?: E E E EDocumento15 pagineWhat Are Enzymes?: E E E EFarhana MuradNessuna valutazione finora
- EnzymesDocumento65 pagineEnzymesnaghalfatih100% (2)
- EnzymesDocumento20 pagineEnzymesRICHARD NESTORNessuna valutazione finora
- BIOCHEM-Lipid Metabolism 1Documento12 pagineBIOCHEM-Lipid Metabolism 1Std Dlshsi100% (1)
- Aspects of Biochemistry LipidsDocumento25 pagineAspects of Biochemistry LipidsLavinia LaviaNessuna valutazione finora
- Carbohydrate MetabolismDocumento83 pagineCarbohydrate Metabolismapi-3826907100% (5)
- CarbohydratesDocumento44 pagineCarbohydratesApril Jean Cahoy100% (3)
- Psychology KMTCDocumento250 paginePsychology KMTCMercy AdeolaNessuna valutazione finora
- Cellular Respiration PDFDocumento10 pagineCellular Respiration PDFAnonymous HXLczq3Nessuna valutazione finora
- Bugg - Introduction To Enzyme and Coenzyme Chemistry 2nd EdDocumento303 pagineBugg - Introduction To Enzyme and Coenzyme Chemistry 2nd EdAlaa13100% (1)
- MitosisDocumento15 pagineMitosisJorge MurieteNessuna valutazione finora
- Biochemistry Lecture 4 Monosaccharides 2Documento48 pagineBiochemistry Lecture 4 Monosaccharides 2Professor Rakesh Sharma Biochemistry Lectures100% (1)
- 1.04 Biochemistry Trans - EnzymesDocumento11 pagine1.04 Biochemistry Trans - EnzymesApril AramNessuna valutazione finora
- Regulation of Gene ExpressionDocumento20 pagineRegulation of Gene ExpressionrainabtNessuna valutazione finora
- Difference Between Alpha and Beta GlucoseDocumento3 pagineDifference Between Alpha and Beta GlucoseLeyla MajundaNessuna valutazione finora
- Biochemistry of Kidneys and UrineDocumento18 pagineBiochemistry of Kidneys and UrineAndrias PutriNessuna valutazione finora
- Chemistry of Lipids: DR Rakesh Sharma, Professor North DMC Medical College, DelhiDocumento50 pagineChemistry of Lipids: DR Rakesh Sharma, Professor North DMC Medical College, DelhiProfessor Rakesh Sharma Biochemistry Lectures100% (1)
- Bacterial MetabolismDocumento46 pagineBacterial MetabolismMakarios S. YousefNessuna valutazione finora
- Micrology and Parasitology ReviewerDocumento44 pagineMicrology and Parasitology Reviewermarjourie cadampog100% (1)
- Biochemistry For Medical Laboratory Science Course Outline 2018Documento4 pagineBiochemistry For Medical Laboratory Science Course Outline 2018Vincent Cuyuca100% (1)
- Classification of Living OrganismsDocumento37 pagineClassification of Living OrganismsSweeetMimiNessuna valutazione finora
- EnzymesDocumento33 pagineEnzymesMary Sutingco100% (1)
- Transport Across Cell MembraneDocumento38 pagineTransport Across Cell MembraneViswadeep DasNessuna valutazione finora
- Cell Cycle Review KeyDocumento4 pagineCell Cycle Review KeyaikaterinaNessuna valutazione finora
- Carbohydrates BiochemistryDocumento14 pagineCarbohydrates Biochemistrylady100% (1)
- Biochemistry PowerpointDocumento14 pagineBiochemistry PowerpointLelaNessuna valutazione finora
- Enzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationDocumento35 pagineEnzymes: A Protein With Catalytic Properties Due To Its Power of Specific ActivationAkash SinghNessuna valutazione finora
- Post Lab Amino Acids and ProteinsDocumento85 paginePost Lab Amino Acids and ProteinsELAINE MARIEJOY MANGAOILNessuna valutazione finora
- 6 - Clinical Biochemistry - 1001411Documento14 pagine6 - Clinical Biochemistry - 1001411bsmallahNessuna valutazione finora
- CarbohydratesDocumento21 pagineCarbohydratesidkmyname100% (5)
- Biochemistry - Chapter 17 Dr. Bongon: Glycolysis & Oxidation of PyruvateDocumento6 pagineBiochemistry - Chapter 17 Dr. Bongon: Glycolysis & Oxidation of PyruvateChaze WaldenNessuna valutazione finora
- 5enzymes and Vitamins PDFDocumento48 pagine5enzymes and Vitamins PDFRomelyn AngelNessuna valutazione finora
- Introduction To Biochemistry: Ms. Jirehkriza G. Suganob General Biology 1Documento27 pagineIntroduction To Biochemistry: Ms. Jirehkriza G. Suganob General Biology 1Jcob SntosNessuna valutazione finora
- Biochemistry LipidsDocumento62 pagineBiochemistry LipidsDianne Joy100% (1)
- Carbohydrate MetabolismDocumento7 pagineCarbohydrate Metabolismcalista osdonNessuna valutazione finora
- LipidsDocumento65 pagineLipidsharisNessuna valutazione finora
- Amino Acid Metabolism NotesDocumento8 pagineAmino Acid Metabolism Notessean100% (2)
- Mic180 - Chapter 5 - Carbohydrate - EditedDocumento75 pagineMic180 - Chapter 5 - Carbohydrate - EditedfarNessuna valutazione finora
- Specificity of EnzymesDocumento14 pagineSpecificity of EnzymesChester James PeñarubiaNessuna valutazione finora
- Biochemistry NotesDocumento25 pagineBiochemistry NotesSelvaArockiam100% (7)
- Chapter 1 - Intorduction To BiochemistryDocumento26 pagineChapter 1 - Intorduction To BiochemistryAnonymous cCCibZnQ1Nessuna valutazione finora
- Lipids BiochemistryDocumento63 pagineLipids Biochemistryanneca100% (1)
- CARBOHYDRATESDocumento31 pagineCARBOHYDRATESCheelsy P. ButronNessuna valutazione finora
- Lab 4 Microbial MetabolismDocumento21 pagineLab 4 Microbial MetabolismEzgi TanılNessuna valutazione finora
- CHPT 4 ENZYMES Lecture Notes (Teacher)Documento6 pagineCHPT 4 ENZYMES Lecture Notes (Teacher)api-3728508100% (3)
- CarbohydratesDocumento156 pagineCarbohydrateseiddnewNessuna valutazione finora
- ProteinsDocumento70 pagineProteinsDianne Joy67% (3)
- Cell Membrane TransportDocumento75 pagineCell Membrane Transportvinay chaudhary100% (1)
- Channels, Carriers, and Pumps: An Introduction to Membrane TransportDa EverandChannels, Carriers, and Pumps: An Introduction to Membrane TransportNessuna valutazione finora
- A Level Organic Chemistry Mini TestDocumento2 pagineA Level Organic Chemistry Mini TestVytheeshwaran VedagiriNessuna valutazione finora
- Fat Malabsorption SyndromeDocumento4 pagineFat Malabsorption SyndromeVytheeshwaran VedagiriNessuna valutazione finora
- Adrenocortical InsufficiencyDocumento5 pagineAdrenocortical InsufficiencyVytheeshwaran VedagiriNessuna valutazione finora
- Copper Deficiency AnemiaDocumento4 pagineCopper Deficiency AnemiaVytheeshwaran VedagiriNessuna valutazione finora
- Type II HypersensitivityDocumento8 pagineType II HypersensitivityVytheeshwaran VedagiriNessuna valutazione finora
- Cushing's SyndromeDocumento4 pagineCushing's SyndromeVytheeshwaran VedagiriNessuna valutazione finora
- LAMARCKISM - Rejected Fixity Proposed ADocumento2 pagineLAMARCKISM - Rejected Fixity Proposed AVytheeshwaran VedagiriNessuna valutazione finora
- GCSE Lower Tier Chemistry TestDocumento5 pagineGCSE Lower Tier Chemistry TestVytheeshwaran VedagiriNessuna valutazione finora
- Hormones SummaryDocumento3 pagineHormones SummaryVytheeshwaran VedagiriNessuna valutazione finora
- Key Stage III Biology Question PaperDocumento6 pagineKey Stage III Biology Question PaperVytheeshwaran VedagiriNessuna valutazione finora
- Dispersal and Fair TestDocumento3 pagineDispersal and Fair TestVytheeshwaran VedagiriNessuna valutazione finora
- Viruses & BacteriophagesDocumento3 pagineViruses & BacteriophagesVytheeshwaran Vedagiri100% (7)
- KS3 TestDocumento6 pagineKS3 TestVytheeshwaran Vedagiri80% (5)
- Island Bio GeographyDocumento2 pagineIsland Bio GeographyVytheeshwaran VedagiriNessuna valutazione finora
- Diabetes MellitusDocumento60 pagineDiabetes MellitusVytheeshwaran Vedagiri96% (26)
- Minitest in Formula and EquationsDocumento2 pagineMinitest in Formula and EquationsVytheeshwaran VedagiriNessuna valutazione finora
- LAMARCKISM - Rejected Fixity Proposed ADocumento2 pagineLAMARCKISM - Rejected Fixity Proposed AVytheeshwaran VedagiriNessuna valutazione finora
- Metabolism inDocumento1 paginaMetabolism inVytheeshwaran VedagiriNessuna valutazione finora
- Clinical Enzymology: V. VytheeshwaranDocumento40 pagineClinical Enzymology: V. VytheeshwaranVytheeshwaran Vedagiri100% (4)
- Candidate GI HormonesDocumento2 pagineCandidate GI HormonesVytheeshwaran Vedagiri100% (2)
- Nucleus - Morphology and FunctionsDocumento25 pagineNucleus - Morphology and FunctionsVytheeshwaran Vedagiri96% (24)
- Bacterial GrowthDocumento5 pagineBacterial GrowthVytheeshwaran Vedagiri100% (20)
- Ventricles of The Brain and CSFDocumento22 pagineVentricles of The Brain and CSFVytheeshwaran Vedagiri100% (9)
- Energetics TestDocumento3 pagineEnergetics TestVytheeshwaran Vedagiri100% (1)
- Unconditional Love of A MotherDocumento3 pagineUnconditional Love of A MotherVytheeshwaran Vedagiri100% (11)
- Interesting Q&A in Immunology - Part IIDocumento26 pagineInteresting Q&A in Immunology - Part IIVytheeshwaran Vedagiri90% (10)
- Golden Words of Sri RamakrishnaDocumento3 pagineGolden Words of Sri RamakrishnaVytheeshwaran Vedagiri100% (13)
- Interesting Q&A in Immunology - Part IDocumento24 pagineInteresting Q&A in Immunology - Part IVytheeshwaran Vedagiri100% (9)
- 11 Sample Papers Physical Education 2020 English Medium Set 1Documento13 pagine11 Sample Papers Physical Education 2020 English Medium Set 1luv kushwahNessuna valutazione finora
- Implan Koklea Dan Fungsi Pendengaran (English Version)Documento29 pagineImplan Koklea Dan Fungsi Pendengaran (English Version)Julita MelisaNessuna valutazione finora
- Patient ManagementDocumento101 paginePatient ManagementRhutvi ViraniNessuna valutazione finora
- Anatomy, Head and Neck, Temporomandibular Joint - StatPearls - NCBI BookshelfDocumento8 pagineAnatomy, Head and Neck, Temporomandibular Joint - StatPearls - NCBI BookshelfDrishti ShahNessuna valutazione finora
- Jurassic Park 1993Documento2 pagineJurassic Park 1993DIVaGNessuna valutazione finora
- Volkan GurelDocumento5 pagineVolkan Gurelapi-392209800Nessuna valutazione finora
- Cerebellum: John H. Martin, Ph.D. Center For Neurobiology & Behavior Columbia UniversityDocumento21 pagineCerebellum: John H. Martin, Ph.D. Center For Neurobiology & Behavior Columbia UniversitySasikala MohanNessuna valutazione finora
- 1 - UK Lamuno Plus BrochureDocumento2 pagine1 - UK Lamuno Plus BrochurePhyo WaiNessuna valutazione finora
- Goby Shirmp MutualismDocumento20 pagineGoby Shirmp MutualismKhandoker Raisul AzadNessuna valutazione finora
- Guia de Algas Bocas Del ToroDocumento71 pagineGuia de Algas Bocas Del ToroNohemi PaolaNessuna valutazione finora
- LFSC Class Test GRD 10 2023Documento4 pagineLFSC Class Test GRD 10 2023gugumatlanatoNessuna valutazione finora
- Connect (4) New QuestionDocumento18 pagineConnect (4) New QuestionGeorge ElsabaNessuna valutazione finora
- GPB 321 Crop Improvement Manual FinalDocumento71 pagineGPB 321 Crop Improvement Manual FinalSultan AhmadNessuna valutazione finora
- Stella MIN - 3.2 BioNinja WorksheetDocumento2 pagineStella MIN - 3.2 BioNinja WorksheetStella Min Eun SuNessuna valutazione finora
- Draft Update Review Artikel Farmakologi Bawang Dayak 040423Documento29 pagineDraft Update Review Artikel Farmakologi Bawang Dayak 040423Zuliar PermanaNessuna valutazione finora
- Erickson's Psycho-Social Theory of DevelopmentDocumento29 pagineErickson's Psycho-Social Theory of Developmentexodo loverNessuna valutazione finora
- Uniit 9 399-408 DepEd-IPDocumento10 pagineUniit 9 399-408 DepEd-IPpeucogcoNessuna valutazione finora
- Heredity and Genetics PowerPointDocumento55 pagineHeredity and Genetics PowerPointShanelle BernalNessuna valutazione finora
- Understanding Immuno-Oncology - The Interplay Between TheDocumento9 pagineUnderstanding Immuno-Oncology - The Interplay Between Thenathalylara050590Nessuna valutazione finora
- His T PathologyDocumento23 pagineHis T PathologygauravNessuna valutazione finora
- Class 8 Winter Break Home-WorkDocumento8 pagineClass 8 Winter Break Home-Workrani bloriaNessuna valutazione finora
- Big Six PDFDocumento4 pagineBig Six PDFmotibaNessuna valutazione finora
- GenBio 1 Lesson 1 ATP and ADP CycleDocumento2 pagineGenBio 1 Lesson 1 ATP and ADP CycleLlaban MunicaNessuna valutazione finora
- Pediatric RehabDocumento32 paginePediatric RehabRainy DaysNessuna valutazione finora
- Bionic EyeDocumento1 paginaBionic EyesubhasishcNessuna valutazione finora