Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Transcription and Translation Exercise VER2 - SolutionDocumento3 pagineTranscription and Translation Exercise VER2 - SolutionMatin Ahmad Khan100% (1)
- Extraintestinal Amebiasis': Scess Was PresentDocumento3 pagineExtraintestinal Amebiasis': Scess Was PresentMaithreyanNandhakumarNessuna valutazione finora
- Fate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyDocumento8 pagineFate of Entamoeba Histolytica During Establishment of Amoebic Liver Abscess Analyzed by Quantitative Radioimaging and HistologyMaithreyanNandhakumarNessuna valutazione finora
- Antifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2Documento93 pagineAntifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2MaithreyanNandhakumarNessuna valutazione finora
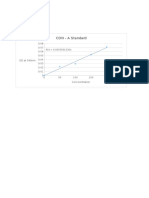
- CON - A Standard: OD at 540nmDocumento10 pagineCON - A Standard: OD at 540nmMaithreyanNandhakumarNessuna valutazione finora
- Sanger SequencingDocumento4 pagineSanger SequencingMilka RahmanNessuna valutazione finora
- HomeostasisDocumento8 pagineHomeostasisLely SharmaNessuna valutazione finora
- Name: TF Name:: LS1a Fall 2014 Lab 3 In-Lab ActivityDocumento4 pagineName: TF Name:: LS1a Fall 2014 Lab 3 In-Lab ActivityThysianNessuna valutazione finora
- The Bacterial RecA Protein and Recombinationlal Dna Repair of Stalled Replication ForksDocumento32 pagineThe Bacterial RecA Protein and Recombinationlal Dna Repair of Stalled Replication ForksHaru SahaNessuna valutazione finora
- CDNA Lab Report - Docx2Documento4 pagineCDNA Lab Report - Docx2cxs5278100% (1)
- Finale Lesson PlanDocumento11 pagineFinale Lesson PlanJayson SorianoNessuna valutazione finora
- QRT PCR Kursus BiomolDocumento40 pagineQRT PCR Kursus BiomolEndang SusilowatiNessuna valutazione finora
- 02 Hormones PDFDocumento8 pagine02 Hormones PDFKaram SaadNessuna valutazione finora
- Protein EngineeringDocumento32 pagineProtein Engineeringmukkucha100% (3)
- Cytogenetics Lesson 2 PrelimDocumento74 pagineCytogenetics Lesson 2 PrelimSinining, Aera Kerl CharmNessuna valutazione finora
- Cell Membrane TransportDocumento3 pagineCell Membrane TransportHarry TeasdaleNessuna valutazione finora
- The Revolution of The CRISPR-Cas9 System EbookDocumento146 pagineThe Revolution of The CRISPR-Cas9 System EbookJose Miguel Fierro BustosNessuna valutazione finora
- Lipids BiochemistryDocumento63 pagineLipids Biochemistryanneca100% (1)
- FSE-Sequencing TechnologyDocumento22 pagineFSE-Sequencing TechnologyYosep GunawanNessuna valutazione finora
- Bio Lab ReportDocumento10 pagineBio Lab Reportapi-313910257Nessuna valutazione finora
- 1841846791Documento600 pagine1841846791Mohammed Hussein100% (1)
- Platinum Taq DNA PolymeraseDocumento2 paginePlatinum Taq DNA PolymeraseAndré OliveiraNessuna valutazione finora
- Daehan College of Business and Technology: Road 20 Sitio Siwang, Nagtinig, Barangay San Juan, Tatay, RizalDocumento2 pagineDaehan College of Business and Technology: Road 20 Sitio Siwang, Nagtinig, Barangay San Juan, Tatay, Rizalangelo llenesNessuna valutazione finora
- Lecture 4 Technical Aspects of Cell Biology 2Documento21 pagineLecture 4 Technical Aspects of Cell Biology 2Jana Moataz Mohamed G02Nessuna valutazione finora
- Lecture 7 Protein Folding: James Chou BCMP201 Spring 2008Documento40 pagineLecture 7 Protein Folding: James Chou BCMP201 Spring 2008KaramSoftNessuna valutazione finora
- G10 Protein SynthesisDocumento41 pagineG10 Protein SynthesisChinsyn FeiNessuna valutazione finora
- nsb1200 1121Documento4 paginensb1200 1121Nicolas RuizNessuna valutazione finora
- L18 - Animal and Plant VirusesDocumento30 pagineL18 - Animal and Plant VirusesM Arfat YameenNessuna valutazione finora
- Molecular Docking 2 - FINALDocumento38 pagineMolecular Docking 2 - FINALvershaparchaNessuna valutazione finora
- Alternative SplicingDocumento25 pagineAlternative Splicingtendril123Nessuna valutazione finora
- Karolin Luger - Nucleosomes: Structure and FunctionDocumento8 pagineKarolin Luger - Nucleosomes: Structure and FunctionDopameNessuna valutazione finora
- Glycolysis Ch.14Documento51 pagineGlycolysis Ch.14Yousef KhallafNessuna valutazione finora
- Protein Function: Burton E. TroppDocumento23 pagineProtein Function: Burton E. TroppLyn JodNessuna valutazione finora
- 3 Transcription-From - DNA - To - RNADocumento7 pagine3 Transcription-From - DNA - To - RNACHIARA ANDREINA ALFARO PURISACANessuna valutazione finora