Potrebbero piacerti anche
- Nematode Infections in Humans, France: Trichostrongylus ColubriformisDocumento2 pagineNematode Infections in Humans, France: Trichostrongylus ColubriformisBeverly FGNessuna valutazione finora
- ErosikkkDocumento20 pagineErosikkkYusrinabilla -Nessuna valutazione finora
- Salmonella Infection of Gallbladder Epithelial Cells Drives Local Inflammation and Injury in A Model of Acute Typhoid FeverDocumento38 pagineSalmonella Infection of Gallbladder Epithelial Cells Drives Local Inflammation and Injury in A Model of Acute Typhoid FeverAgussalimNessuna valutazione finora
- Feline Gastrointestinal Eosinofílica Fibroplasia EsclerosanteDocumento8 pagineFeline Gastrointestinal Eosinofílica Fibroplasia EsclerosanteCarlos Alberto Chaves VelasquezNessuna valutazione finora
- J Immunol 2007 Rydström 5789 801Documento14 pagineJ Immunol 2007 Rydström 5789 801susana1616Nessuna valutazione finora
- Jun Zou, Nathan ShankarDocumento11 pagineJun Zou, Nathan ShankarHQ HQNessuna valutazione finora
- Helmintos TisularesDocumento8 pagineHelmintos TisularesValentina Arenas RNessuna valutazione finora
- Antibiotic Resistance Profile of Staphylococci From Clinical Sources Recovered From InfantsDocumento7 pagineAntibiotic Resistance Profile of Staphylococci From Clinical Sources Recovered From InfantsCornius TerranovaNessuna valutazione finora
- Developmental and Comparative Immunology: Ying Zhang, Hui Sun, Huilin Zhao, Xingxing Chen, Jiaojiao Li, Boqing LiDocumento6 pagineDevelopmental and Comparative Immunology: Ying Zhang, Hui Sun, Huilin Zhao, Xingxing Chen, Jiaojiao Li, Boqing LiMohammad Farhan AdityaNessuna valutazione finora
- Emetine Produce Entamoeba Histolytica Death by IndDocumento6 pagineEmetine Produce Entamoeba Histolytica Death by IndJota CarlosNessuna valutazione finora
- Feline Panleukopenia Virus Infection in Various Species From HungaryDocumento9 pagineFeline Panleukopenia Virus Infection in Various Species From HungaryAlonSultanNessuna valutazione finora
- Jir 409Documento10 pagineJir 409utkarsh PromotiomNessuna valutazione finora
- A Mouse Model of Recrudescence ofDocumento4 pagineA Mouse Model of Recrudescence ofPaulo CzarnewskiNessuna valutazione finora
- Vet Pathol 2013 Delaney 172 6 DolphinDocumento6 pagineVet Pathol 2013 Delaney 172 6 DolphinKhairul Ihsan Pengusaha MudaNessuna valutazione finora
- 2014 Article 47 PDFDocumento7 pagine2014 Article 47 PDFalif bagusNessuna valutazione finora
- Fimmu 11 571816Documento20 pagineFimmu 11 571816Ricardo GomezNessuna valutazione finora
- Comparative Methods For Demonstration: Study of Tissue Culture of Toxoplasma GondiiDocumento4 pagineComparative Methods For Demonstration: Study of Tissue Culture of Toxoplasma GondiiSalim JufriNessuna valutazione finora
- Immunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient MouseDocumento5 pagineImmunohistochemical Characterization of A Renal Nephroblastoma in A Trp53-Mutant and Prolyl Isomerase 1-Deficient Mouselily rogaNessuna valutazione finora
- Causal Agents: AmoebiasisDocumento5 pagineCausal Agents: Amoebiasisadinda sandyaNessuna valutazione finora
- Immunomodulation in The Canine Endometrium by UteropathogenicDocumento17 pagineImmunomodulation in The Canine Endometrium by UteropathogenicAndreea Blanco VeraNessuna valutazione finora
- Jced 7 E656Documento4 pagineJced 7 E656María José VázquezNessuna valutazione finora
- CHAPTER 2-ParasitDocumento109 pagineCHAPTER 2-Parasitalery ahreallyNessuna valutazione finora
- Jurnal Patogenesis AmoebiasisDocumento12 pagineJurnal Patogenesis AmoebiasisAngga NuralamNessuna valutazione finora
- Amoebiasis NewDocumento27 pagineAmoebiasis NewMutiaNessuna valutazione finora
- 1 s2.0 S0002944018305698 MainDocumento9 pagine1 s2.0 S0002944018305698 MainFrank Barreiro SanchezNessuna valutazione finora
- Diagnosis and Management of Amebiasis: State-Of-The-Art Clinical ArticleDocumento9 pagineDiagnosis and Management of Amebiasis: State-Of-The-Art Clinical ArticleSiscaSelviaNessuna valutazione finora
- Green DKK, 2015Documento14 pagineGreen DKK, 2015Nabila NatasyaNessuna valutazione finora
- Influence of Acute-Phase Parasite Load On PathologyDocumento11 pagineInfluence of Acute-Phase Parasite Load On PathologymclimacoNessuna valutazione finora
- 1 2 BRDocumento3 pagine1 2 BRrahmani bagherNessuna valutazione finora
- Cat L PDFDocumento11 pagineCat L PDFFlaviu TabaranNessuna valutazione finora
- Intestinal Protozoa Amoebae & Ciliates: ObjectivesDocumento5 pagineIntestinal Protozoa Amoebae & Ciliates: ObjectivesmicroperadeniyaNessuna valutazione finora
- Anthrax Lethal Toxin Disrupts Intestinal Barrier Function and Causes Systemic Infections With Enteric Bacteria 2012Documento10 pagineAnthrax Lethal Toxin Disrupts Intestinal Barrier Function and Causes Systemic Infections With Enteric Bacteria 2012fer123wizNessuna valutazione finora
- Recent Buzz in Malaria Research: The FEBS Journal, Editorial Office, Cambridge, UKDocumento4 pagineRecent Buzz in Malaria Research: The FEBS Journal, Editorial Office, Cambridge, UKVerlyanita SeptiariniNessuna valutazione finora
- Acute HepatotoxityDocumento11 pagineAcute HepatotoxityThially BragaNessuna valutazione finora
- Lancet - Septic Shock (Seminars)Documento16 pagineLancet - Septic Shock (Seminars)williamsbarriosNessuna valutazione finora
- Medmicro 32 2 135Documento7 pagineMedmicro 32 2 135vinty permatasariNessuna valutazione finora
- Artículo 2 - Fisiopatología de La PielonefritisDocumento17 pagineArtículo 2 - Fisiopatología de La PielonefritisCAROLINA CHUMACERO BERMEONessuna valutazione finora
- The Malaria Cell Atlas A Comprehensive RDocumento50 pagineThe Malaria Cell Atlas A Comprehensive R9868838836ankNessuna valutazione finora
- Gamberini Et AlDocumento9 pagineGamberini Et AlDeboraXiningNessuna valutazione finora
- Gut Bacteriome and Metabolome of Ascaris Lumbricoides in PatientsDocumento13 pagineGut Bacteriome and Metabolome of Ascaris Lumbricoides in PatientsprofnutrialecuriNessuna valutazione finora
- Wang 2015Documento7 pagineWang 2015Sebastian MilesNessuna valutazione finora
- 2021 Genome-Wide Association Study of Resistance To MTBDocumento18 pagine2021 Genome-Wide Association Study of Resistance To MTBAntonio Jafar GarciaNessuna valutazione finora
- Aeruginosa Strain PAO1 Is Regulated by Quorum Sensing andDocumento15 pagineAeruginosa Strain PAO1 Is Regulated by Quorum Sensing andSarah KKCNessuna valutazione finora
- Entamoba 1Documento15 pagineEntamoba 1nisaaintanNessuna valutazione finora
- Amebiasis: Review ArticleDocumento9 pagineAmebiasis: Review ArticleJouffrey Itaar MadridistaNessuna valutazione finora
- Hongos PerroDocumento4 pagineHongos PerroHelen Dellepiane GilNessuna valutazione finora
- Invasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BDocumento5 pagineInvasive Epithelial Mesothelioma in A Dog: F. R, B. B, K. R, E. S, D. BjamesyuNessuna valutazione finora
- Clinical, Morphological and Immunohistochemical Characterization of Cutaneous Lymphocytosis in 23 Cats (Pages 3-12)Documento10 pagineClinical, Morphological and Immunohistochemical Characterization of Cutaneous Lymphocytosis in 23 Cats (Pages 3-12)jenNessuna valutazione finora
- 1 s2.0 S1286457912001918 MainDocumento8 pagine1 s2.0 S1286457912001918 Mainabznaim420Nessuna valutazione finora
- Petton 2021 Frontiers Pacific MANUSCRIT2Documento26 paginePetton 2021 Frontiers Pacific MANUSCRIT2AmadorRevillaNessuna valutazione finora
- J Infect Dis.-2000-Hang-1738-48Documento11 pagineJ Infect Dis.-2000-Hang-1738-48Anonymous hXd6PJlNessuna valutazione finora
- E. Coli TITIDocumento6 pagineE. Coli TITIiggyNessuna valutazione finora
- Toxoplasma Gondii: Regulates ICAM-1 Mediated Monocyte Adhesion To TrophoblastsDocumento7 pagineToxoplasma Gondii: Regulates ICAM-1 Mediated Monocyte Adhesion To TrophoblastsCarlos Javier Castro CavadiaNessuna valutazione finora
- 423 432 PDFDocumento10 pagine423 432 PDFaldiansyahraufNessuna valutazione finora
- Original Article: A Guide To Histomorphological Evaluation of Intestinal Inflammation in Mouse ModelsDocumento21 pagineOriginal Article: A Guide To Histomorphological Evaluation of Intestinal Inflammation in Mouse ModelsOUR STORYNessuna valutazione finora
- Apoptosis in Natural Rabies Virus Infection in DogsDocumento5 pagineApoptosis in Natural Rabies Virus Infection in DogspapiipiiNessuna valutazione finora
- Candidatus Odyssella Thessalonicensis ' Gen. Nov., Sp. Nov., An Obligate Intracellular Parasite of Acanthamoeba SpeciesDocumento10 pagineCandidatus Odyssella Thessalonicensis ' Gen. Nov., Sp. Nov., An Obligate Intracellular Parasite of Acanthamoeba SpeciessomasushmaNessuna valutazione finora
- Parasitos en ReptilesDocumento13 pagineParasitos en ReptilesEduardo Pella100% (1)
- A Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPDocumento22 pagineA Quantitative PCR (Taqman) Assay For Pathogenic: Leptospira SPPVigneshwaran RavishankarNessuna valutazione finora
- Aikat1979 PDFDocumento5 pagineAikat1979 PDFMaithreyanNandhakumarNessuna valutazione finora
- Alternative Telomere RepaiirDocumento16 pagineAlternative Telomere RepaiirMaithreyanNandhakumarNessuna valutazione finora
- The Present and Future Role of Microfluidics PDFDocumento9 pagineThe Present and Future Role of Microfluidics PDFMaithreyanNandhakumarNessuna valutazione finora
- ThreeChannel PDFDocumento8 pagineThreeChannel PDFMaithreyanNandhakumarNessuna valutazione finora
- Extraintestinal Amebiasis': Scess Was PresentDocumento3 pagineExtraintestinal Amebiasis': Scess Was PresentMaithreyanNandhakumarNessuna valutazione finora
- Mechanotransduction PDFDocumento8 pagineMechanotransduction PDFMaithreyanNandhakumarNessuna valutazione finora
- 3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningDocumento11 pagine3microfluidic Co-Culture System For Cancer Migratory Analysis and Anti-Metastatic Drugs ScreeningMaithreyanNandhakumarNessuna valutazione finora
- Current Concept Same Bias IsDocumento10 pagineCurrent Concept Same Bias IsMaithreyanNandhakumarNessuna valutazione finora
- 4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerDocumento18 pagine4A Microfluidic 3D in Vitro Model For Specificity of Breast CancerMaithreyanNandhakumarNessuna valutazione finora
- 5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsDocumento29 pagine5microfluidic Assay For Simultaneous Culture of Multiple Cell Types On Surfaces or Within HydrogelsMaithreyanNandhakumarNessuna valutazione finora
- 2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfDocumento25 pagine2investigating Metastasis Using in Vitro Platforms - Madame Curie Bioscience Database - NCBI BookshelfMaithreyanNandhakumarNessuna valutazione finora
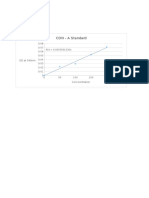
- CON - A Standard: OD at 540nmDocumento10 pagineCON - A Standard: OD at 540nmMaithreyanNandhakumarNessuna valutazione finora
- 1microfluidic ModellingDocumento22 pagine1microfluidic ModellingMaithreyanNandhakumarNessuna valutazione finora
- ReferencesDocumento4 pagineReferencesMaithreyanNandhakumarNessuna valutazione finora
- Introduction To Flow Cytometry - A Learning GuideDocumento54 pagineIntroduction To Flow Cytometry - A Learning GuideFrank FontaineNessuna valutazione finora
- Antifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2Documento93 pagineAntifreeze Proteins Enhance Survival of Cells in Cryopreservation - Substituting DMSO With RmAFP#1 in Cryopreservation of Cells 1.2MaithreyanNandhakumarNessuna valutazione finora
- .Documento33 pagine.3bdullah al3yuniNessuna valutazione finora
- Factors Affecting Wound HealingDocumento11 pagineFactors Affecting Wound HealingFredy Rodeardo MaringgaNessuna valutazione finora
- Pathology FinalADocumento72 paginePathology FinalAvaegmundig100% (1)
- What You Should Know About LipogemsDocumento4 pagineWhat You Should Know About LipogemsEmma KalmanNessuna valutazione finora
- Interpretation of The HemogramDocumento5 pagineInterpretation of The HemogramMargarita MujicaNessuna valutazione finora
- COVID Thrombosis: Mitochondria, The Endothelial Glycocalyx and Platelets Are KeyDocumento10 pagineCOVID Thrombosis: Mitochondria, The Endothelial Glycocalyx and Platelets Are KeyMercedes BouterNessuna valutazione finora
- DR - Bambang-Damage Control OrthopaedicsDocumento37 pagineDR - Bambang-Damage Control OrthopaedicsbadliinaaNessuna valutazione finora
- Pathophysiology of CHF Secondary To RHDDocumento89 paginePathophysiology of CHF Secondary To RHDMira MariantiNessuna valutazione finora
- Shar-Pei Dogs Hypocobalaminemia PDFDocumento10 pagineShar-Pei Dogs Hypocobalaminemia PDFFlávia UchôaNessuna valutazione finora
- Biomarcadores em IBDDocumento7 pagineBiomarcadores em IBDAndressa RigoNessuna valutazione finora
- Burns 3Documento31 pagineBurns 3Reann LeeNessuna valutazione finora
- Aktu Syllabus Bpharm 2nd Sem PDFDocumento21 pagineAktu Syllabus Bpharm 2nd Sem PDFAnkit Kumar100% (1)
- Nutrigenomics and CancerDocumento9 pagineNutrigenomics and CancerAbyatar Hugo BaskoroNessuna valutazione finora
- Pathology Notes (Class Notes)Documento106 paginePathology Notes (Class Notes)Mustafa Kamal Bangash100% (1)
- Protein Energy Wasting What Is It and What Can We Do To Prevent ItDocumento8 pagineProtein Energy Wasting What Is It and What Can We Do To Prevent ItLú VillalobosNessuna valutazione finora
- 1 Fundamentals of Nursing PracticeDocumento27 pagine1 Fundamentals of Nursing PracticeJoseph Manglallan100% (1)
- Ncma219 Lec FinalDocumento45 pagineNcma219 Lec FinalangeliqueggonzalesNessuna valutazione finora
- Novak Et Al 2011 AllergyDocumento10 pagineNovak Et Al 2011 AllergyArturo VeraNessuna valutazione finora
- Lecture 3 - Immunological System (K.Dhaliwal 2019)Documento62 pagineLecture 3 - Immunological System (K.Dhaliwal 2019)brittany AcheampongNessuna valutazione finora
- Culture of Animal Cells: Fig. 1.2. Tissue Culture ApplicationsDocumento1 paginaCulture of Animal Cells: Fig. 1.2. Tissue Culture ApplicationsCatalina ColoradoNessuna valutazione finora
- SF Journal of Biotechnology and Biomedical EngineeringDocumento5 pagineSF Journal of Biotechnology and Biomedical EngineeringFAUZANNessuna valutazione finora
- Peripatetic Cardiomyo - Genetics To ManagementDocumento11 paginePeripatetic Cardiomyo - Genetics To Managementanna67890Nessuna valutazione finora
- MelkiorDocumento5 pagineMelkiorFiLiOeiNessuna valutazione finora
- Systemic - Sclerosis Nature Review 2014Documento22 pagineSystemic - Sclerosis Nature Review 2014CT TANNessuna valutazione finora
- Robbins Chapter 2 DiagramsDocumento20 pagineRobbins Chapter 2 Diagramsjeffaguilar100% (2)
- Gastritis in DogsDocumento2 pagineGastritis in DogsPamela BriceñoNessuna valutazione finora
- Articol2 1Documento6 pagineArticol2 1Smaranda GavrilNessuna valutazione finora
- ProceedingsofROINMED2013 PDFDocumento161 pagineProceedingsofROINMED2013 PDFRazvan GheorgheNessuna valutazione finora
- SerratiopeptidaseDocumento4 pagineSerratiopeptidaseMo KhanNessuna valutazione finora
- Infrared Radiation PDFDocumento10 pagineInfrared Radiation PDFAjay Pal NattNessuna valutazione finora