Potrebbero piacerti anche
- 10 PDFDocumento1 pagina10 PDFChrisNessuna valutazione finora
- EKG PediatrikDocumento3 pagineEKG PediatrikChrisNessuna valutazione finora
- Sinopsis PsikiatriDocumento5 pagineSinopsis PsikiatriChrisNessuna valutazione finora
- MR IcuDocumento42 pagineMR IcuChrisNessuna valutazione finora
- HE Catalogue Jul15 FinalDocumento42 pagineHE Catalogue Jul15 FinalChrisNessuna valutazione finora
- Fear of EyesDocumento16 pagineFear of EyesChrisNessuna valutazione finora
- Melloni's Pocket Medical DictionaryDocumento2.992 pagineMelloni's Pocket Medical DictionaryChrisNessuna valutazione finora
- Week 1 SundayDocumento1 paginaWeek 1 SundaydsadasdsakdjsakljNessuna valutazione finora
- Paranoma ExcisionDocumento1 paginaParanoma ExcisionChrisNessuna valutazione finora
- Week 1 Tuesday Thursday SaturdayDocumento1 paginaWeek 1 Tuesday Thursday SaturdaydsadasdsakdjsakljNessuna valutazione finora
- 1diphtheria Spain Rapid Risk Assessment June 2015Documento8 pagine1diphtheria Spain Rapid Risk Assessment June 2015Francisca BallesterosNessuna valutazione finora
- Anti ArrhythmicdrugsDocumento52 pagineAnti ArrhythmicdrugsChrisNessuna valutazione finora
- Result of Hyperthyroidism: Sumber: Mayo ClinicDocumento9 pagineResult of Hyperthyroidism: Sumber: Mayo ClinicChrisNessuna valutazione finora
- Week 1 Monday Wednesday FridayDocumento1 paginaWeek 1 Monday Wednesday FridaydsadasdsakdjsakljNessuna valutazione finora
- Scoliosis Exercises: Physical Therapy DepartmentDocumento4 pagineScoliosis Exercises: Physical Therapy DepartmentIfa10371% (7)
- Why So Many Scoliosis Patients Are Using The Spinecor Brace and The Schroth Method of Exercises Together (PDF Reference)Documento3 pagineWhy So Many Scoliosis Patients Are Using The Spinecor Brace and The Schroth Method of Exercises Together (PDF Reference)ChrisNessuna valutazione finora
- Scoliosis Exercises: Physical Therapy DepartmentDocumento4 pagineScoliosis Exercises: Physical Therapy DepartmentIfa10371% (7)
- Exercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)Documento16 pagineExercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)ChrisNessuna valutazione finora
- Exercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)Documento16 pagineExercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)ChrisNessuna valutazione finora
- Exercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)Documento16 pagineExercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)ChrisNessuna valutazione finora
- Pentalaksanaan RBDocumento84 paginePentalaksanaan RBFara Sakina RahmaNessuna valutazione finora
- Exercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)Documento16 pagineExercises For Adolescent Idiopathic Scoliosis (Protocol) (PDF Reference)ChrisNessuna valutazione finora
- The Way People Dress Reveals Their Personality and Taste.: GoldDocumento3 pagineThe Way People Dress Reveals Their Personality and Taste.: GoldChrisNessuna valutazione finora
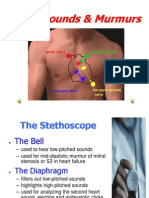
- Heart Sounds and MurmursDocumento38 pagineHeart Sounds and MurmursLaura Moise100% (5)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)