Potrebbero piacerti anche
- Curt Hoys 2013Documento12 pagineCurt Hoys 2013broken22Nessuna valutazione finora
- Part 1 - US-Grade 4-BiochemistryDocumento34 paginePart 1 - US-Grade 4-BiochemistryFarah Bashar Al-RawachyNessuna valutazione finora
- Renaltubularacidosis: Robert Todd Alexander,, Martin BitzanDocumento23 pagineRenaltubularacidosis: Robert Todd Alexander,, Martin BitzanRicardo CastañedaNessuna valutazione finora
- 6-Drugs Acting On The Renal System - 2Documento65 pagine6-Drugs Acting On The Renal System - 2Hamid Hussain HamidNessuna valutazione finora
- Inorganic and Organic Anion Transport by Insect Renal EpitheliaDocumento13 pagineInorganic and Organic Anion Transport by Insect Renal EpitheliaCLPHtheoryNessuna valutazione finora
- Lec 14 - Kidney PDFDocumento16 pagineLec 14 - Kidney PDFrajeshNessuna valutazione finora
- GFR measurement using inulin clearanceDocumento12 pagineGFR measurement using inulin clearanceVanathy KarunamoorthyNessuna valutazione finora
- 2013 Renal Tubular AcidosisDocumento13 pagine2013 Renal Tubular AcidosisLUIS FRANCISCO GUERRERO MARTÍNEZNessuna valutazione finora
- J of Gastro and Hepatol - 2001 - Strazzabosco - Pathophysiology of The Intrahepatic Biliary EpitheliumDocumento10 pagineJ of Gastro and Hepatol - 2001 - Strazzabosco - Pathophysiology of The Intrahepatic Biliary EpitheliumJosé LeãoNessuna valutazione finora
- Intestinal Ion Transport and The Pathophysiology of DiarrheaDocumento13 pagineIntestinal Ion Transport and The Pathophysiology of Diarrheajuanfraflores9777Nessuna valutazione finora
- Renal Physiology: Tubular ProcessingDocumento6 pagineRenal Physiology: Tubular ProcessingUloko ChristopherNessuna valutazione finora
- Obat Pada Ginjal: Prof M.A.Widodo PHDDocumento53 pagineObat Pada Ginjal: Prof M.A.Widodo PHDPriiya Ashiwini100% (1)
- Acid-Base and Potassium HomeostasisDocumento8 pagineAcid-Base and Potassium HomeostasisVeronica WongNessuna valutazione finora
- Renal System 2: Transport Properties of Nephron SegmentsDocumento5 pagineRenal System 2: Transport Properties of Nephron SegmentsSherida GibbsNessuna valutazione finora
- Renal Extra Notes: Total Body Water (TBW) 0.6 X Body WeightDocumento3 pagineRenal Extra Notes: Total Body Water (TBW) 0.6 X Body WeightKhaing26Nessuna valutazione finora
- General Principles of Tubular ReabsorptionDocumento54 pagineGeneral Principles of Tubular ReabsorptionRalph JacintoNessuna valutazione finora
- Disorder of Potassium HemostasisDocumento9 pagineDisorder of Potassium HemostasisGevanski Hermanov MaturbongsNessuna valutazione finora
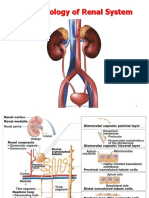
- Pharmacology of Renal SystemDocumento125 paginePharmacology of Renal SystemBirhanu GetaNessuna valutazione finora
- Compare and Contrast Ionic Movements Across Renal and Digestive Tract EpitheliaDocumento2 pagineCompare and Contrast Ionic Movements Across Renal and Digestive Tract EpitheliaDhruvalNessuna valutazione finora
- Tubular ReabsorptionDocumento42 pagineTubular Reabsorptionmina mounirNessuna valutazione finora
- Intestinal Ion Transport and The Pathophysiology of Diarrhea - PMCDocumento22 pagineIntestinal Ion Transport and The Pathophysiology of Diarrhea - PMCtanatswa courtney matingoNessuna valutazione finora
- Lecture 20 Notes 2018Documento6 pagineLecture 20 Notes 2018Omed ZarifiNessuna valutazione finora
- Physiology - BSDocumento14 paginePhysiology - BSkep1313Nessuna valutazione finora
- Kidney Function and Metabolism RegulationDocumento21 pagineKidney Function and Metabolism RegulationLuis OñaNessuna valutazione finora
- Sodium, Water and Potassium: Michael D. PenneyDocumento39 pagineSodium, Water and Potassium: Michael D. PenneyChristianus LeonardNessuna valutazione finora
- 101 OHara PDFDocumento11 pagine101 OHara PDFAntonio Lopez HernandezNessuna valutazione finora
- Mechanisms of Disease: Review ArticleDocumento11 pagineMechanisms of Disease: Review ArticleDámaris Barrueto MezaNessuna valutazione finora
- Distal Convoluted Tubules, Counter Current Mechanism, MicturitionDocumento94 pagineDistal Convoluted Tubules, Counter Current Mechanism, MicturitionSarah BirechNessuna valutazione finora
- Distal Tubule Balance and Tubuloglomerular Feedback-Group 2Documento42 pagineDistal Tubule Balance and Tubuloglomerular Feedback-Group 2Chuleha SethNessuna valutazione finora
- Tubular Reabsorption and SecretionDocumento29 pagineTubular Reabsorption and SecretionenriNessuna valutazione finora
- L3 +L4 Renal System 2023Documento31 pagineL3 +L4 Renal System 2023mizare29gNessuna valutazione finora
- Drugs Affecting Renal FunctionDocumento52 pagineDrugs Affecting Renal Functionliakot prantoNessuna valutazione finora
- Mechanism of Filteration of Urine 5Documento34 pagineMechanism of Filteration of Urine 5Akshay Pratap SinghNessuna valutazione finora
- Pathogenesis, Diagnosis and Management of Hyperkalemia PDFDocumento9 paginePathogenesis, Diagnosis and Management of Hyperkalemia PDFJbarrianNessuna valutazione finora
- Chapter 18 DIURETICSDocumento14 pagineChapter 18 DIURETICSAlexandra AlexaNessuna valutazione finora
- Acidosis TubularDocumento5 pagineAcidosis TubularLuisFranciscoPaniaguaLopezNessuna valutazione finora
- UrinaryDocumento87 pagineUrinaryJohn Paolo Dalida SalazarNessuna valutazione finora
- Chloride The Queen of ElectrolytesDocumento9 pagineChloride The Queen of ElectrolytesYancy Erazo DoradoNessuna valutazione finora
- Molecular Cell Biology Lodish 7th Edition Solutions ManualDocumento8 pagineMolecular Cell Biology Lodish 7th Edition Solutions ManualKennethHendersonyecg100% (39)
- Fluid Electrolytes and Nutrition Physiological and Clinical AspectsDocumento14 pagineFluid Electrolytes and Nutrition Physiological and Clinical AspectsPaulynhiita FashyOnNessuna valutazione finora
- Theory UrinalysisDocumento23 pagineTheory UrinalysisAnggi Dyah AristiNessuna valutazione finora
- Topik 3Documento29 pagineTopik 3AnnisNessuna valutazione finora
- Mechanism of Urine FormationDocumento5 pagineMechanism of Urine FormationovacuteNessuna valutazione finora
- Ion TransportersDocumento2 pagineIon TransportersfilipemedicinavrNessuna valutazione finora
- Liquidos y Electrolitos 2Documento4 pagineLiquidos y Electrolitos 2Jorge MBNessuna valutazione finora
- Chapter 19 - The Urinary System: Fluid and Electrolyte BalanceDocumento28 pagineChapter 19 - The Urinary System: Fluid and Electrolyte BalanceSara SabraNessuna valutazione finora
- Solution Manual For Molecular Cell Biology 8th Edition Harvey Lodish Arnold Berk Chris A Kaiser Monty Krieger Anthony Bretscher Hidde Ploegh Angelika Amon Kelsey C Martin 33Documento5 pagineSolution Manual For Molecular Cell Biology 8th Edition Harvey Lodish Arnold Berk Chris A Kaiser Monty Krieger Anthony Bretscher Hidde Ploegh Angelika Amon Kelsey C Martin 33PhilipWoodpsen100% (37)
- Sir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsDocumento15 pagineSir David Cuthbertson Medal Lecture - Fluid, Electrolytes and Nutrition: Physiological and Clinical AspectsThinh VinhNessuna valutazione finora
- QuestionsDocumento4 pagineQuestionsA GoswamiNessuna valutazione finora
- Hepatocellular Transport in Acquired Cholestasis: New Insights Into Functional, Regulatory and Therapeutic AspectsDocumento22 pagineHepatocellular Transport in Acquired Cholestasis: New Insights Into Functional, Regulatory and Therapeutic AspectsdimbrutNessuna valutazione finora
- Electrolyte Disorders: Understanding Sodium and Potassium HomeostasisDocumento14 pagineElectrolyte Disorders: Understanding Sodium and Potassium HomeostasisBayu Budi SukocoNessuna valutazione finora
- SX FalconiDocumento15 pagineSX FalconiJesús QuirarteNessuna valutazione finora
- Ranal SystemDocumento35 pagineRanal SystemAdil IrshadNessuna valutazione finora
- Biochemistry of Kidneys and UrineDocumento18 pagineBiochemistry of Kidneys and UrineAndrias PutriNessuna valutazione finora
- Thyroid Hormone Deficiency Alters Expression of AcDocumento13 pagineThyroid Hormone Deficiency Alters Expression of AcNatty NigussieNessuna valutazione finora
- Cellular Respiration PDFDocumento15 pagineCellular Respiration PDFJovicaNessuna valutazione finora
- Chapter 24. The Kidney.: 1. The Structure and Function of The NephronDocumento8 pagineChapter 24. The Kidney.: 1. The Structure and Function of The NephronjeskevandiemenNessuna valutazione finora
- Bases Da Fisiopatologia Da Diarreia (The Pathophysiology of Diarrhea)Documento18 pagineBases Da Fisiopatologia Da Diarreia (The Pathophysiology of Diarrhea)MujiartiNessuna valutazione finora
- Hisam Nurokhman.a (201902010094)Documento22 pagineHisam Nurokhman.a (201902010094)Hisam hoboNessuna valutazione finora
- Renal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledDa EverandRenal Reflections: Exploring the Marvels of Nephrology: The Intricate Ballet of Kidney Functions UnveiledNessuna valutazione finora
- Process Industry Practices Electrical: Pip Eleha01 Engineering Guide For Determining Electrical Area ClassificationDocumento21 pagineProcess Industry Practices Electrical: Pip Eleha01 Engineering Guide For Determining Electrical Area Classificationhrol31060Nessuna valutazione finora
- Standard Steel Doors and FramesDocumento11 pagineStandard Steel Doors and Frameshrol31060100% (1)
- Sewer DetailsDocumento18 pagineSewer Detailshrol31060100% (1)
- Saddle AnalysisDocumento17 pagineSaddle AnalysisAntonio Perez ZornozaNessuna valutazione finora
- Partially Full Pipe Flow Calculations Using Excel Spreadsheets 7-18-14Documento28 paginePartially Full Pipe Flow Calculations Using Excel Spreadsheets 7-18-14hrol310600% (1)
- Partially Full Pipe Flow Calculations Using Excel Spreadsheets 7-18-14Documento28 paginePartially Full Pipe Flow Calculations Using Excel Spreadsheets 7-18-14hrol310600% (1)
- PIP ARC01015 Architectural and Building Utilities Design CriteriaDocumento39 paginePIP ARC01015 Architectural and Building Utilities Design Criteriahrol31060100% (1)
- Civil Design Basis - B PDFDocumento45 pagineCivil Design Basis - B PDFhrol31060Nessuna valutazione finora
- Blast Resistant Design With Structural SteelDocumento6 pagineBlast Resistant Design With Structural SteelJavier Flores100% (1)
- Steel Design - LRFD Bolted Connections TutorialDocumento4 pagineSteel Design - LRFD Bolted Connections TutorialFernando PizarroNessuna valutazione finora
- Instantaneous Center of Rotation Method PDFDocumento3 pagineInstantaneous Center of Rotation Method PDFhrol31060Nessuna valutazione finora
- Horizontal Vessel Foundation DesignDocumento42 pagineHorizontal Vessel Foundation Designhrol31060100% (1)
- 2017 NASCC Intro 15ed ManualDocumento25 pagine2017 NASCC Intro 15ed Manualhrol31060Nessuna valutazione finora
- Analysis of Shear Lag in Steel Angle ConnectorsDocumento34 pagineAnalysis of Shear Lag in Steel Angle ConnectorsGonzalo ContrerasNessuna valutazione finora
- 30 Good Rules for Cost-Effective Connection DesignDocumento4 pagine30 Good Rules for Cost-Effective Connection Designมานะ จรูญพงษ์ศักดิ์Nessuna valutazione finora
- Chronic Kidney DiseaseDocumento11 pagineChronic Kidney Diseasehrol31060Nessuna valutazione finora
- 0602 1 472Documento6 pagine0602 1 472hrol31060Nessuna valutazione finora
- Blast Resistant Building Design CriteriaDocumento51 pagineBlast Resistant Building Design Criteriahrol31060100% (2)
- 28 93 Cemento Portland EspecificacionesDocumento12 pagine28 93 Cemento Portland Especificacionesrmejias_sNessuna valutazione finora
- Vertical Vessel Foundation DesignDocumento33 pagineVertical Vessel Foundation Designhrol31060100% (2)
- F436Documento6 pagineF436hrol31060Nessuna valutazione finora
- DB Sections and BarsDocumento1 paginaDB Sections and Barshrol31060Nessuna valutazione finora
- 1272 FullDocumento10 pagine1272 Fullhrol31060Nessuna valutazione finora
- Insanity Brazil Butt Lift Hybrid WorkoutDocumento11 pagineInsanity Brazil Butt Lift Hybrid WorkoutGil Medina-FloresNessuna valutazione finora
- Chronic Kidney DiseaseDocumento11 pagineChronic Kidney Diseasehrol31060Nessuna valutazione finora
- 1461 FullDocumento9 pagine1461 Fullhrol31060Nessuna valutazione finora
- Beam Column Base Plate DesignDocumento10 pagineBeam Column Base Plate DesignSPUD1100% (3)
- A Test in Phonetics 500 Questions and Answers On English Pronunciation - Dr. B. SiertsemaDocumento91 pagineA Test in Phonetics 500 Questions and Answers On English Pronunciation - Dr. B. SiertsemaTrasgu Viajero67% (3)
- Am J Kidney Dis 2014 Oct 64 (4) 510Documento24 pagineAm J Kidney Dis 2014 Oct 64 (4) 510hrol31060Nessuna valutazione finora
- Biology Chapter 21 Sylvia Mader ModifiedDocumento29 pagineBiology Chapter 21 Sylvia Mader ModifiedabdallahNessuna valutazione finora
- Chapter 10 Mendilan and Genetics Study GuideDocumento16 pagineChapter 10 Mendilan and Genetics Study GuideKing AllanNessuna valutazione finora
- The First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Documento5 pagineThe First Comparative Examination 2021-22: Class XII (ISC) BIOLOGY Paper-1 (Theory)Chinmay SinghNessuna valutazione finora
- 13 Miller Chap 4b LectureDocumento21 pagine13 Miller Chap 4b LectureGerone Tolentino AtienzaNessuna valutazione finora
- DNA and RNA - Genetic MaterialDocumento10 pagineDNA and RNA - Genetic MaterialsahanaNessuna valutazione finora
- 2022 2024 Syllabus - RemovedDocumento35 pagine2022 2024 Syllabus - Removednj639830Nessuna valutazione finora
- Viral Classification: DNA VirusesDocumento10 pagineViral Classification: DNA Virusess.khan9211rediffmail.comNessuna valutazione finora
- Eoc Biology Rview Packet 2012-2013Documento48 pagineEoc Biology Rview Packet 2012-2013api-282682755Nessuna valutazione finora
- Genetic Information and Protein SynthesisDocumento22 pagineGenetic Information and Protein Synthesisjason balmasNessuna valutazione finora
- NEET Question Paper 2022 ExamDocumento74 pagineNEET Question Paper 2022 ExamSudhirNessuna valutazione finora
- Mirna 146 and Its Association With Autoimmune DseasesDocumento7 pagineMirna 146 and Its Association With Autoimmune DseasesIJAR JOURNALNessuna valutazione finora
- Antiviral Agents Chapter Provides Overview of Classification and BiochemistryDocumento26 pagineAntiviral Agents Chapter Provides Overview of Classification and BiochemistryEmily Larissa Saldanha PereiraNessuna valutazione finora
- BIOMOLECULES Proteins and Nucleic AcidsDocumento23 pagineBIOMOLECULES Proteins and Nucleic Acidsjia aganaNessuna valutazione finora
- Brazilian Society For Food and Nutrition Position Statement - Nutrigenetic TestsDocumento12 pagineBrazilian Society For Food and Nutrition Position Statement - Nutrigenetic TestsDaniel Carvalho de AraújoNessuna valutazione finora
- RnaiDocumento13 pagineRnaiIrah Jane BadeNessuna valutazione finora
- Biology Grade 11 Dna & RnaDocumento9 pagineBiology Grade 11 Dna & RnaBread UniverseNessuna valutazione finora
- 8.12 RNA Transcription WorksheetDocumento2 pagine8.12 RNA Transcription WorksheetAmor Reyes EngNessuna valutazione finora
- DNA - The Double Helix, Coloring WorksheetDocumento4 pagineDNA - The Double Helix, Coloring WorksheetRob MullerNessuna valutazione finora
- Pomegranate - Botany, Postharvest Treatment, Biochemical Composition and Health Effects (2014) PDFDocumento188 paginePomegranate - Botany, Postharvest Treatment, Biochemical Composition and Health Effects (2014) PDFAndré Campelo100% (2)
- Regulation of Gene ExpressionDocumento5 pagineRegulation of Gene ExpressionHanumat SinghNessuna valutazione finora
- Mcqs ImbDocumento23 pagineMcqs ImbMuvawala Belekia AbelNessuna valutazione finora
- MATH3353 NotesDocumento100 pagineMATH3353 NotesOwen CameronNessuna valutazione finora
- BCHN+213 2023 T2memoDocumento5 pagineBCHN+213 2023 T2memowhoeversaidthis1Nessuna valutazione finora
- Genetic Analysis An Integrative Approach 1st Edition Sanders Test BankDocumento12 pagineGenetic Analysis An Integrative Approach 1st Edition Sanders Test BankHeatherAllencejrx100% (16)
- Otot 2Documento106 pagineOtot 2anita parwatiNessuna valutazione finora
- SB 3Documento8 pagineSB 3diyasanjeev13Nessuna valutazione finora
- Claribel Ria MaeDocumento19 pagineClaribel Ria MaeGLENN MENDOZANessuna valutazione finora
- Cell 1981 Cech PDFDocumento10 pagineCell 1981 Cech PDFSpellkingNessuna valutazione finora
- 3.molecular Diagnosis CourseDocumento120 pagine3.molecular Diagnosis CourseSorin LazarNessuna valutazione finora
- DNA, RNA and Protein Synthesis ExplainedDocumento12 pagineDNA, RNA and Protein Synthesis ExplainedrorNessuna valutazione finora