Potrebbero piacerti anche
- Temática - Ciências da Vida - Relação e CoordenaçãoDa EverandTemática - Ciências da Vida - Relação e CoordenaçãoNessuna valutazione finora
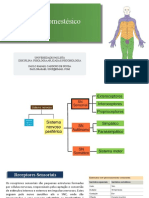
- 05 - Sistema SomestésicoDocumento27 pagine05 - Sistema SomestésicoPaulo RafaelNessuna valutazione finora
- Resumo Sistema SensorialDocumento11 pagineResumo Sistema SensorialMaya LumaNessuna valutazione finora
- Sistema SensorialDocumento27 pagineSistema SensorialAmanda100% (2)
- Sistema SOMATOSSENSITIVODocumento8 pagineSistema SOMATOSSENSITIVOVictor MarquesNessuna valutazione finora
- SomestesiaDocumento9 pagineSomestesiaJuliana PissolatoNessuna valutazione finora
- Fisiologia Dos Órgãos Dos SentidosDocumento38 pagineFisiologia Dos Órgãos Dos SentidosJonathan MazaiaNessuna valutazione finora
- 2 RevisoSistema Nervoso Parte IIDocumento28 pagine2 RevisoSistema Nervoso Parte IIsamuelwerickrNessuna valutazione finora
- Sensibilidade SomáticaDocumento22 pagineSensibilidade SomáticaAntonio Zandre Moreira Lopes100% (1)
- Resumo - Fisiologia NeuromuscularDocumento5 pagineResumo - Fisiologia NeuromuscularMarina MarquesNessuna valutazione finora
- Sistema Somato-SensorialDocumento33 pagineSistema Somato-SensorialCleandro Ribeiro Do ValleNessuna valutazione finora
- Neurofisiologia Sensitiva e MotoraDocumento8 pagineNeurofisiologia Sensitiva e MotoraJaqueline Dias100% (1)
- Sistema SensorialDocumento11 pagineSistema SensorialPedro Henrique Souza MatsumuraNessuna valutazione finora
- Sensibilidade SomáticaDocumento46 pagineSensibilidade SomáticaVandecir SilvaNessuna valutazione finora
- MÓDULO 2 Estimulação TegumentarDocumento41 pagineMÓDULO 2 Estimulação TegumentarmelnicNessuna valutazione finora
- Estudo Dirigido Receptores Sensoriais e Circuitos NeuronaisDocumento3 pagineEstudo Dirigido Receptores Sensoriais e Circuitos NeuronaisWillian GonzagaNessuna valutazione finora
- Vias AscendentesDocumento9 pagineVias AscendentesLucianaNessuna valutazione finora
- NeurofisiologiaDocumento37 pagineNeurofisiologiaingridcanaverdeNessuna valutazione finora
- TatoDocumento12 pagineTatoCamila MendesNessuna valutazione finora
- SomestesiaDocumento4 pagineSomestesiaNoemi SantosNessuna valutazione finora
- Sistema Nervoso Sensorial SomaticoDocumento105 pagineSistema Nervoso Sensorial SomaticoAna Larissa Castro100% (3)
- Receptores SensoriaisDocumento45 pagineReceptores SensoriaisJuliana CostaNessuna valutazione finora
- 6 Texto - Os Sentidos SomáticosDocumento4 pagine6 Texto - Os Sentidos SomáticosCoordenação BiomedicinaNessuna valutazione finora
- Artigo 1 TBL 1Documento34 pagineArtigo 1 TBL 1Ian JhemesNessuna valutazione finora
- Sistema SensorialDocumento83 pagineSistema SensorialElvis Rheinschmitt100% (2)
- FisiologiaDocumento2 pagineFisiologiaIzabele AmandaNessuna valutazione finora
- Neurofisiologia Aula 3Documento60 pagineNeurofisiologia Aula 3Maria Clara Figueira BarbosaNessuna valutazione finora
- ApostilaDocumento14 pagineApostilaAnderson LimaNessuna valutazione finora
- #Resumo Sensações#Documento9 pagine#Resumo Sensações#Monitoriadefisio100% (4)
- Fisio - P1 PDFDocumento9 pagineFisio - P1 PDFDaniele VitoriaNessuna valutazione finora
- Apostila Taping Estético Editada 050419Documento18 pagineApostila Taping Estético Editada 050419Priscila Biggi100% (1)
- Fisiologia Sensorial 24Documento31 pagineFisiologia Sensorial 24Ester MelloNessuna valutazione finora
- Sistema NervosoDocumento21 pagineSistema NervosoJéssica QueirozNessuna valutazione finora
- Sensação Somática e DorDocumento45 pagineSensação Somática e DorBrunaNessuna valutazione finora
- SinapseDocumento17 pagineSinapseJuliana Vilhena Evangelista da CostaNessuna valutazione finora
- Sist SensorialDocumento3 pagineSist SensorialIsadora GnbaNessuna valutazione finora
- Aula 2 Parte I - Nocicepção e TemperaturaDocumento15 pagineAula 2 Parte I - Nocicepção e TemperaturaPatrícia SilvaNessuna valutazione finora
- Somestesia ArtigoDocumento6 pagineSomestesia ArtigoFúlvia NicollyNessuna valutazione finora
- Questões Fisio Miv 8Documento7 pagineQuestões Fisio Miv 8Amanda ANessuna valutazione finora
- Reflexos EspinhaisDocumento29 pagineReflexos EspinhaisAnderson Sartor PedroniNessuna valutazione finora
- Neuro P1-F1Documento9 pagineNeuro P1-F1Ana Luiza SoutoNessuna valutazione finora
- MOTRICIDADEDocumento12 pagineMOTRICIDADEirisreaisNessuna valutazione finora
- 6 Sistema Nervoso SensorialDocumento40 pagine6 Sistema Nervoso SensorialWander Junior100% (1)
- Abordagem SensorialDocumento7 pagineAbordagem SensorialReinaldo Dos AnjosNessuna valutazione finora
- Fisiologia - 04 - Receptores Sensoriais e CircuitosDocumento38 pagineFisiologia - 04 - Receptores Sensoriais e CircuitosMatheusNessuna valutazione finora
- Nociceptores e Hiperalgesia.Documento32 pagineNociceptores e Hiperalgesia.Kedma VitóriaNessuna valutazione finora
- Teste para Mapeamento Das Sensações CutâneasDocumento5 pagineTeste para Mapeamento Das Sensações CutâneasPedro Lino100% (1)
- Sistema Nervoso Sensorial Monitoria Aula 3 PDFDocumento7 pagineSistema Nervoso Sensorial Monitoria Aula 3 PDFSabrynna LimaNessuna valutazione finora
- FISIOLOGIADocumento22 pagineFISIOLOGIAAna PaulaNessuna valutazione finora
- Relatório Fisiologia Sistema SensorialDocumento17 pagineRelatório Fisiologia Sistema SensorialArquimeniaNessuna valutazione finora
- Aula 03 - Fisiologia SensorialDocumento85 pagineAula 03 - Fisiologia SensorialTadeu Alves100% (1)
- FisiologiaDocumento15 pagineFisiologiaKaroline GilbertNessuna valutazione finora
- 06 SomestesiaDocumento18 pagine06 SomestesiaOdilon Calian0% (1)
- Tutoria 3 - TatoDocumento12 pagineTutoria 3 - Tatopaulo josé da silvaNessuna valutazione finora
- Sistema SomassensorialDocumento5 pagineSistema SomassensorialAnuarite CanoquelaNessuna valutazione finora
- PORTF3Documento61 paginePORTF3Nicole SantosNessuna valutazione finora
- Caderno de Estudos Práticos em NeuroanatomiaDa EverandCaderno de Estudos Práticos em NeuroanatomiaNessuna valutazione finora
- Transfusão de HemocomponentesDocumento2 pagineTransfusão de HemocomponentesBreno SouzaNessuna valutazione finora
- Anemia MegaloblásticaDocumento3 pagineAnemia MegaloblásticaBreno SouzaNessuna valutazione finora
- Mieloma MúltiploDocumento2 pagineMieloma MúltiploBreno SouzaNessuna valutazione finora
- LeishmanioseDocumento1 paginaLeishmanioseBreno SouzaNessuna valutazione finora
- Linfoadenomegalias e LinfomasDocumento3 pagineLinfoadenomegalias e LinfomasBreno SouzaNessuna valutazione finora
- Anemia FerroprivaDocumento4 pagineAnemia FerroprivaBreno SouzaNessuna valutazione finora
- AnemiasDocumento2 pagineAnemiasBreno SouzaNessuna valutazione finora
- AnemiasDocumento2 pagineAnemiasBreno SouzaNessuna valutazione finora
- Insuficiência Renal Aguda e Métodos DialíticosDocumento3 pagineInsuficiência Renal Aguda e Métodos DialíticosBreno SouzaNessuna valutazione finora
- Infecção Do Trato Urinário Definição e EtiologiaDocumento2 pagineInfecção Do Trato Urinário Definição e EtiologiaBreno SouzaNessuna valutazione finora
- GlomerulopatiasDocumento3 pagineGlomerulopatiasBreno SouzaNessuna valutazione finora
- Insuficiência Renal Crônica e Transplante RenalDocumento4 pagineInsuficiência Renal Crônica e Transplante RenalBreno SouzaNessuna valutazione finora
- Aparelho RespiratórioDocumento6 pagineAparelho RespiratórioBreno SouzaNessuna valutazione finora
- Anemia FerroprivaDocumento4 pagineAnemia FerroprivaBreno SouzaNessuna valutazione finora
- GametogêneseDocumento8 pagineGametogêneseBreno SouzaNessuna valutazione finora
- Tecido ÓsseoDocumento6 pagineTecido ÓsseoaeadaNessuna valutazione finora
- 3° Prova de Embriologia Sistema Nervoso e SensorialDocumento10 pagine3° Prova de Embriologia Sistema Nervoso e SensorialAnonymous 8VKtmd2NQ9Nessuna valutazione finora
- Resumo de FarmacologiaDocumento21 pagineResumo de FarmacologiaBreno Souza100% (2)
- DorsoDocumento12 pagineDorsoBreno SouzaNessuna valutazione finora
- Histologia Sistema Reprodutor FemininoDocumento8 pagineHistologia Sistema Reprodutor FemininoBreno SouzaNessuna valutazione finora
- Tecido SanguíneoDocumento4 pagineTecido SanguíneoBreno SouzaNessuna valutazione finora
- Fisica QuimicaDocumento17 pagineFisica QuimicaTobiasKnightNessuna valutazione finora
- Exp8 Apresentacao 20Documento18 pagineExp8 Apresentacao 20anitadiogoNessuna valutazione finora
- Histologia Da OrelhaDocumento5 pagineHistologia Da OrelhaMaria Paula RangelNessuna valutazione finora
- Apostila Sonoplastia PDFDocumento36 pagineApostila Sonoplastia PDFVladimir Borges100% (1)
- ESTESIOLOGIADocumento22 pagineESTESIOLOGIAInês VenâncioNessuna valutazione finora
- Neuroanatomia e Neurologia - Recurso Pedagógico - ExercíciosDocumento25 pagineNeuroanatomia e Neurologia - Recurso Pedagógico - Exercíciosrichard santosNessuna valutazione finora
- Auriculo Chinesa EadDocumento79 pagineAuriculo Chinesa EadLuciano JonackNessuna valutazione finora
- APOSTILA UNIP PPB - Processos Psicológicos BásicosDocumento117 pagineAPOSTILA UNIP PPB - Processos Psicológicos BásicosAléxia Vega100% (7)
- A Língua de Sinais Como Recurso para A Terapia FonoaudiológicaDocumento32 pagineA Língua de Sinais Como Recurso para A Terapia FonoaudiológicaCarla VidalNessuna valutazione finora
- Plano de Aula Ed InfantilDocumento21 paginePlano de Aula Ed InfantillucianeNessuna valutazione finora
- Gate 02 - Contato (2 Parte)Documento207 pagineGate 02 - Contato (2 Parte)tiulau.minhapaginaNessuna valutazione finora
- Monstro Do LagoDocumento12 pagineMonstro Do LagoDuda100% (2)
- Aula03 Acustica COMENTADADocumento6 pagineAula03 Acustica COMENTADAprifabiojorge2Nessuna valutazione finora
- Apostila Módulo I - LibrasDocumento52 pagineApostila Módulo I - Librasyasmin.adrienne98Nessuna valutazione finora
- Pontos de NogierdocDocumento8 paginePontos de NogierdocBruno Souza de MarchiNessuna valutazione finora
- Slide Aparelho AuditivoDocumento9 pagineSlide Aparelho AuditivoJéssica SantanaNessuna valutazione finora
- Escala Bohlen-Pierce: Concepção MacrotonalDocumento80 pagineEscala Bohlen-Pierce: Concepção MacrotonalFlavio Aoun SapienzaNessuna valutazione finora
- Artigo de Divulgação CientificaDocumento2 pagineArtigo de Divulgação CientificaFatima OliveiraNessuna valutazione finora
- 3 - ROTEIRO MORFOFUNCIONAL - Neurofisiologia Do Equilibrio, Sistema Vestibular, Sentidos Correlatos Com QuedaDocumento8 pagine3 - ROTEIRO MORFOFUNCIONAL - Neurofisiologia Do Equilibrio, Sistema Vestibular, Sentidos Correlatos Com QuedaINGRID MILENA MARQUES DE ANDRADENessuna valutazione finora
- Samir Gerges - HeadseatDocumento3 pagineSamir Gerges - HeadseatGustavo AlcântaraNessuna valutazione finora
- Easy - Programa de Treinamento Auditivo r3Documento2 pagineEasy - Programa de Treinamento Auditivo r3Luiz Cysne0% (1)
- Medicina Do TráfegoDocumento24 pagineMedicina Do Tráfego4kdjwgmb9nNessuna valutazione finora
- 024 - Mergulho Riscos À Saúde Dos MergulhadoresDocumento74 pagine024 - Mergulho Riscos À Saúde Dos MergulhadoresclaudioNessuna valutazione finora
- Deixe Ela Gozar 10 Vezes Antes de VocãDocumento160 pagineDeixe Ela Gozar 10 Vezes Antes de VocãGilmar Botelho100% (2)
- Sistemas de TelecomunicaçõesDocumento123 pagineSistemas de TelecomunicaçõesThiago MaiaNessuna valutazione finora
- Aula Pratica - Sistema Auditivo e Vestibular - 2024Documento4 pagineAula Pratica - Sistema Auditivo e Vestibular - 2024vivisaggiomoNessuna valutazione finora
- AUT0278 Aula 03 - Som, Ouvido e Percepção SonoraDocumento36 pagineAUT0278 Aula 03 - Som, Ouvido e Percepção SonoraBaixar ApostilaNessuna valutazione finora
- Gary Carpenter PDFDocumento3 pagineGary Carpenter PDFCasa Do cachorro quenteNessuna valutazione finora
- Cheklist Anamnese e Exame FísicoDocumento7 pagineCheklist Anamnese e Exame FísicoIzael Silva100% (2)
- Apostila de TosaDocumento41 pagineApostila de Tosaluiz_cdias100% (2)