Potrebbero piacerti anche
- Trade Offs in Liquid AlternativesDocumento11 pagineTrade Offs in Liquid AlternativesChris HeasmanNessuna valutazione finora
- WalkinBlues Open GDocumento8 pagineWalkinBlues Open GChris HeasmanNessuna valutazione finora
- Emerging Markets Hedge Funds-Capturing Alpha in An Inefficient Asset ClassDocumento11 pagineEmerging Markets Hedge Funds-Capturing Alpha in An Inefficient Asset ClassChris HeasmanNessuna valutazione finora
- Alternative Investmetns - Early InvestingDocumento12 pagineAlternative Investmetns - Early InvestingChris HeasmanNessuna valutazione finora
- Mandolin Tab and Notation PDFDocumento1 paginaMandolin Tab and Notation PDFChris HeasmanNessuna valutazione finora
- 6 Line Tab and NotationDocumento1 pagina6 Line Tab and NotationMeltedman De-constructionNessuna valutazione finora
- Akin Gump HF StructureDocumento12 pagineAkin Gump HF StructureChris HeasmanNessuna valutazione finora
- MalaguenaDocumento2 pagineMalaguenaGabriele ZanettiNessuna valutazione finora
- 1993 Duffield Al - FLWR Choice by Honey BeesDocumento7 pagine1993 Duffield Al - FLWR Choice by Honey BeesChris HeasmanNessuna valutazione finora
- Aspects of Foraging in Bees - Apple Pollination, Native Bee Populations, and Honey Bee CommunicationDocumento90 pagineAspects of Foraging in Bees - Apple Pollination, Native Bee Populations, and Honey Bee CommunicationChris HeasmanNessuna valutazione finora
- FoodTrends 2016Documento75 pagineFoodTrends 2016ThuyDuongNessuna valutazione finora
- Emerging Markets Hedge Funds: Capturing Alpha in An Inefficient Asset ClassDocumento11 pagineEmerging Markets Hedge Funds: Capturing Alpha in An Inefficient Asset ClassJason McCoyNessuna valutazione finora
- Nosema Disease: Literature Review and Three Year Survey of BeekeepersDocumento35 pagineNosema Disease: Literature Review and Three Year Survey of BeekeepersChris HeasmanNessuna valutazione finora
- Regional Training Southern 2 Beekeeping Africa Malawi Training enDocumento44 pagineRegional Training Southern 2 Beekeeping Africa Malawi Training enChris HeasmanNessuna valutazione finora
- Antibiotic-Specific Differences in The Response of Staphylococcus Aureus To Treatment With Antimicrobials Combined With Manuka HoneyDocumento9 pagineAntibiotic-Specific Differences in The Response of Staphylococcus Aureus To Treatment With Antimicrobials Combined With Manuka HoneyChris HeasmanNessuna valutazione finora
- Deciphering The Frontier Opportunity - Lazard Perspectives PaperDocumento8 pagineDeciphering The Frontier Opportunity - Lazard Perspectives PaperChris HeasmanNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Ford Focus MK2 Headlight Switch Wiring DiagramDocumento1 paginaFord Focus MK2 Headlight Switch Wiring DiagramAdam TNessuna valutazione finora
- The Intel 8086 / 8088/ 80186 / 80286 / 80386 / 80486 Jump InstructionsDocumento3 pagineThe Intel 8086 / 8088/ 80186 / 80286 / 80386 / 80486 Jump InstructionsalexiouconNessuna valutazione finora
- Intro To MavenDocumento18 pagineIntro To MavenDaniel ReckerthNessuna valutazione finora
- HDO OpeationsDocumento28 pagineHDO OpeationsAtif NadeemNessuna valutazione finora
- Bill of Quantities 16FI0009Documento1 paginaBill of Quantities 16FI0009AJothamChristianNessuna valutazione finora
- 3) Uses and Gratification: 1) The Hypodermic Needle ModelDocumento5 pagine3) Uses and Gratification: 1) The Hypodermic Needle ModelMarikaMcCambridgeNessuna valutazione finora
- ID25bc8b496-2013 Dse English PaperDocumento2 pagineID25bc8b496-2013 Dse English PaperSimpson WainuiNessuna valutazione finora
- Acute Suppurative Otitis MediaDocumento41 pagineAcute Suppurative Otitis Mediarani suwadjiNessuna valutazione finora
- Nat Steel BREGENEPD000379Documento16 pagineNat Steel BREGENEPD000379Batu GajahNessuna valutazione finora
- Consumer Protection ActDocumento34 pagineConsumer Protection ActshikhroxNessuna valutazione finora
- 2015 NSPC Proposed Changes Book PDFDocumento268 pagine2015 NSPC Proposed Changes Book PDFMichelle Pinohermoso JabatNessuna valutazione finora
- Introduction Spreadable Media TtsDocumento22 pagineIntroduction Spreadable Media TtsYanro FerrerNessuna valutazione finora
- Painters Rates PDFDocumento86 paginePainters Rates PDFmanthoexNessuna valutazione finora
- Legrand Price List-01 ST April-2014Documento144 pagineLegrand Price List-01 ST April-2014Umesh SutharNessuna valutazione finora
- Concise Selina Solutions Class 9 Maths Chapter 15 Construction of PolygonsDocumento31 pagineConcise Selina Solutions Class 9 Maths Chapter 15 Construction of Polygonsbhaskar51178Nessuna valutazione finora
- BV DoshiDocumento6 pagineBV DoshiNimNessuna valutazione finora
- Level of Organisation of Protein StructureDocumento18 pagineLevel of Organisation of Protein Structureyinghui94Nessuna valutazione finora
- Pediatric Fever of Unknown Origin: Educational GapDocumento14 paginePediatric Fever of Unknown Origin: Educational GapPiegl-Gulácsy VeraNessuna valutazione finora
- Communication Skill - Time ManagementDocumento18 pagineCommunication Skill - Time ManagementChấn NguyễnNessuna valutazione finora
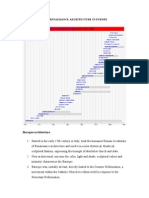
- Post Renaissance Architecture in EuropeDocumento10 paginePost Renaissance Architecture in Europekali_007Nessuna valutazione finora
- 2010 Information ExchangeDocumento15 pagine2010 Information ExchangeAnastasia RotareanuNessuna valutazione finora
- Our Story Needs No Filter by Nagarkar SudeepDocumento153 pagineOur Story Needs No Filter by Nagarkar SudeepKavya SunderNessuna valutazione finora
- 8 X 56 M.-SCH.: Country of Origin: ATDocumento1 pagina8 X 56 M.-SCH.: Country of Origin: ATMohammed SirelkhatimNessuna valutazione finora
- Transactions List: Marilena Constantin RO75BRDE445SV93146784450 RON Marilena ConstantinDocumento12 pagineTransactions List: Marilena Constantin RO75BRDE445SV93146784450 RON Marilena ConstantinConstantin MarilenaNessuna valutazione finora
- ARIIX - Clean - Eating - Easy - Ecipes - For - A - Healthy - Life - Narx PDFDocumento48 pagineARIIX - Clean - Eating - Easy - Ecipes - For - A - Healthy - Life - Narx PDFAnte BaškovićNessuna valutazione finora
- Docket - CDB Batu GajahDocumento1 paginaDocket - CDB Batu Gajahfatin rabiatul adawiyahNessuna valutazione finora
- 2015 Student Handbook 16 January 2015Documento66 pagine2015 Student Handbook 16 January 2015John KhanNessuna valutazione finora
- Tec066 6700 PDFDocumento2 pagineTec066 6700 PDFExclusivo VIPNessuna valutazione finora
- Domesticity and Power in The Early Mughal WorldDocumento17 pagineDomesticity and Power in The Early Mughal WorldUjjwal Gupta100% (1)
- Ethical Hacking IdDocumento24 pagineEthical Hacking IdSilvester Dian Handy PermanaNessuna valutazione finora