Potrebbero piacerti anche
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsDa EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNessuna valutazione finora
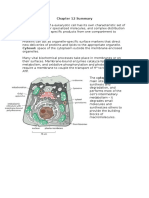
- Chapter 16 SummaryDocumento11 pagineChapter 16 SummaryCharlotteNessuna valutazione finora
- Chapter 15 SummaryDocumento27 pagineChapter 15 SummaryCharlotteNessuna valutazione finora
- Chapter 4 SummaryDocumento8 pagineChapter 4 SummaryCharlotteNessuna valutazione finora
- Biochemistry FinalsDocumento11 pagineBiochemistry FinalsKc CalderonNessuna valutazione finora
- Chapter 20 Solutions Molecular Biology of The CellDocumento15 pagineChapter 20 Solutions Molecular Biology of The CellAkshey BadrinarayanNessuna valutazione finora
- Genome & Personality-2ndDocumento28 pagineGenome & Personality-2ndVrendra Kumar SharmaNessuna valutazione finora
- Amino Acid: Prepared By: Anthonette Ann Alemania-STEM 12Documento28 pagineAmino Acid: Prepared By: Anthonette Ann Alemania-STEM 12Anthonette Alemania100% (1)
- Epidemiology 2 PDFDocumento11 pagineEpidemiology 2 PDFJawad TatlahNessuna valutazione finora
- Medicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryDa EverandMedicinal Chemistry—III: Main Lectures Presented at the Third International Symposium on Medicinal ChemistryP. PratesiNessuna valutazione finora
- Waves As PhysicsDocumento8 pagineWaves As PhysicsRija MughalNessuna valutazione finora
- Chapter 6 SummaryDocumento28 pagineChapter 6 SummaryCharlotteNessuna valutazione finora
- Unit 1: Introduction: Organic InorganicDocumento1 paginaUnit 1: Introduction: Organic InorganicEllah GutierrezNessuna valutazione finora
- Chapter 12 SummaryDocumento7 pagineChapter 12 SummaryCharlotteNessuna valutazione finora
- Histology Notes 2Documento13 pagineHistology Notes 2Lukas BüchlerNessuna valutazione finora
- Review For The 2° Semester Exam Alessandro Mo6a, UVVG, 3 YearDocumento9 pagineReview For The 2° Semester Exam Alessandro Mo6a, UVVG, 3 Yeardjxela89Nessuna valutazione finora
- Cell Signaling IDocumento21 pagineCell Signaling IGuhanNessuna valutazione finora
- Basic Molecular Biology & Biotechnology Training ManualDocumento23 pagineBasic Molecular Biology & Biotechnology Training Manualsameer420Nessuna valutazione finora
- Enzyme Inhibitors Explained: Reversible vs IrreversibleDocumento5 pagineEnzyme Inhibitors Explained: Reversible vs IrreversiblebelabelawNessuna valutazione finora
- Final Exam Study Guide TipsDocumento22 pagineFinal Exam Study Guide Tipsrazgriz1211Nessuna valutazione finora
- IUPAC NomenclatureDocumento17 pagineIUPAC Nomenclaturesurya kant upadhyay100% (3)
- Neural Tube DefectsDocumento12 pagineNeural Tube Defectsdaniel_1592Nessuna valutazione finora
- Epidemiology EpidemiologyDocumento13 pagineEpidemiology EpidemiologyBiratNagarNessuna valutazione finora
- Signal Transduction Mechanisms and Cellular ResponsesDocumento39 pagineSignal Transduction Mechanisms and Cellular ResponsesBellony Sanders100% (1)
- Cell Biology Review: Blood Smear and Connective TissueDocumento15 pagineCell Biology Review: Blood Smear and Connective TissueGabriel ParasNessuna valutazione finora
- Test 4 Essay Questoin Meiosis and GametogenesisDocumento2 pagineTest 4 Essay Questoin Meiosis and GametogenesisInactiveAccount80% (5)
- 05 Connective TissueDocumento43 pagine05 Connective TissuetanzirineNessuna valutazione finora
- Chapter 13 SummaryDocumento5 pagineChapter 13 SummaryCharlotteNessuna valutazione finora
- Musculoskeletal PhysiologyDocumento5 pagineMusculoskeletal PhysiologyKhairul IkhwanNessuna valutazione finora
- 2 - Bones of The Lower LimbDocumento23 pagine2 - Bones of The Lower LimbHamzaNessuna valutazione finora
- MEN1 PPDocumento15 pagineMEN1 PPAaron D. PhoenixNessuna valutazione finora
- Notes About BiologyDocumento81 pagineNotes About BiologyRichard CoffeyNessuna valutazione finora
- Ecg Lab Bio350Documento13 pagineEcg Lab Bio350Robin Lewis CooperNessuna valutazione finora
- Pharynx: Upper Part: Widest - 3.5 CM, Non-Collapsible Middle Part: Narrow Lower End: Narrowest Part of GITDocumento5 paginePharynx: Upper Part: Widest - 3.5 CM, Non-Collapsible Middle Part: Narrow Lower End: Narrowest Part of GITdrpnnreddyNessuna valutazione finora
- Soft Tissue TumourDocumento8 pagineSoft Tissue TumourjslumNessuna valutazione finora
- Exam 1 NotesDocumento54 pagineExam 1 NotesAnonymous If9p21kwZKNessuna valutazione finora
- Summary Molecular Biology of The Cell Chapter 17Documento9 pagineSummary Molecular Biology of The Cell Chapter 17Yi Jun Hu100% (2)
- Bsci201 Anatomy Class NotesDocumento101 pagineBsci201 Anatomy Class NotesSahel UddinNessuna valutazione finora
- Sama Notes Upper LimbDocumento15 pagineSama Notes Upper LimbMaiUyen Kieu50% (2)
- NPTEL – Cellular and Molecular Immunology: Antibodies and AntigensDocumento33 pagineNPTEL – Cellular and Molecular Immunology: Antibodies and AntigensAygul RamankulovaNessuna valutazione finora
- Cell Signalling 1.2 2017Documento61 pagineCell Signalling 1.2 2017PutriNessuna valutazione finora
- Excitation Contraction Coupling: Nandini GoyalDocumento32 pagineExcitation Contraction Coupling: Nandini GoyalNandini GoyalNessuna valutazione finora
- Overview of Cell Cycle by Javali.GDocumento15 pagineOverview of Cell Cycle by Javali.GJavali.GNessuna valutazione finora
- Cranial Nerves DiagramDocumento15 pagineCranial Nerves DiagramSpellkingNessuna valutazione finora
- Module 6 Portfolio PDFDocumento3 pagineModule 6 Portfolio PDFAndrea Celine AurealNessuna valutazione finora
- Intermediate FilamentsDocumento4 pagineIntermediate FilamentsSai SridharNessuna valutazione finora
- Spectrum Classes: Chemical Bonding ExplainedDocumento33 pagineSpectrum Classes: Chemical Bonding ExplainedRANJEET SHARMANessuna valutazione finora
- Dysplastic and Proliferative DisordersDocumento10 pagineDysplastic and Proliferative DisordersYA M AYNessuna valutazione finora
- Histology of GIT Part 1 Esophagus PDFDocumento20 pagineHistology of GIT Part 1 Esophagus PDFJeffrey RamosNessuna valutazione finora
- 04 Mediastinum, HeartDocumento27 pagine04 Mediastinum, HeartShaira Aquino VerzosaNessuna valutazione finora
- Multidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseDocumento12 pagineMultidimensional Analysis of Clinical Symptoms in Patients With Fabry's DiseaseOnon EssayedNessuna valutazione finora
- HistologyDocumento61 pagineHistologysofy_aboshokNessuna valutazione finora
- Cell Structures FinalDocumento16 pagineCell Structures Finalapi-269480689Nessuna valutazione finora
- Osmolarity&tonicitypracticeproblemsfrominternetDocumento14 pagineOsmolarity&tonicitypracticeproblemsfrominternetbsktblbabe88Nessuna valutazione finora
- BASC02 Full PDFDocumento70 pagineBASC02 Full PDFndarubagusNessuna valutazione finora
- Genetic Control of Cell Function and InheritanceDocumento7 pagineGenetic Control of Cell Function and InheritanceSADAVEJO100% (1)
- Physiology. Hrd.Documento549 paginePhysiology. Hrd.Sʌɩĸʌt PʌʋɭNessuna valutazione finora
- Electrochemistry Lect Notes CambridgeDocumento4 pagineElectrochemistry Lect Notes Cambridgeavatar_75Nessuna valutazione finora
- Histology of The Upper GITDocumento43 pagineHistology of The Upper GITBenedict OkotNessuna valutazione finora
- Antibiotic Break Down ChartDocumento4 pagineAntibiotic Break Down ChartBram StokerNessuna valutazione finora
- Chapter 19 SummaryDocumento10 pagineChapter 19 SummaryCharlotteNessuna valutazione finora
- Chapter 8 SummaryDocumento25 pagineChapter 8 SummaryCharlotteNessuna valutazione finora
- Chapter 20 SummaryyDocumento21 pagineChapter 20 SummaryyCharlotteNessuna valutazione finora
- Chapter 17 SummaryDocumento22 pagineChapter 17 SummaryCharlotteNessuna valutazione finora
- Chapter 13 SummaryDocumento5 pagineChapter 13 SummaryCharlotteNessuna valutazione finora
- Chapter 7 SummaryDocumento34 pagineChapter 7 SummaryCharlotteNessuna valutazione finora
- Chapter 3 SummaryDocumento10 pagineChapter 3 SummaryCharlotteNessuna valutazione finora
- Chapter 6 SummaryDocumento28 pagineChapter 6 SummaryCharlotteNessuna valutazione finora
- Chapter 12 SummaryDocumento7 pagineChapter 12 SummaryCharlotteNessuna valutazione finora
- Chapter 2 SummaryDocumento3 pagineChapter 2 SummaryCharlotteNessuna valutazione finora
- Chapter 1 SummaryDocumento4 pagineChapter 1 SummaryCharlotteNessuna valutazione finora
- The Physiology of AgingDocumento19 pagineThe Physiology of AgingHoney TuizaNessuna valutazione finora
- Cadmium, Organ Toxicity and Therapeutic Approaches: A Review On Brain, Kidney and Testis DamageDocumento15 pagineCadmium, Organ Toxicity and Therapeutic Approaches: A Review On Brain, Kidney and Testis DamageRama RaynsfordNessuna valutazione finora
- An Anti Apoptotic HEK293 Cell Line Provides A Robust and High Titer Platform For Transient Protein Expression in BioreactorsDocumento11 pagineAn Anti Apoptotic HEK293 Cell Line Provides A Robust and High Titer Platform For Transient Protein Expression in BioreactorsAtrocitus RedNessuna valutazione finora
- How Cancer Cells Remodel Lipid Metabolism: Strategies Targeting Transcription FactorsDocumento19 pagineHow Cancer Cells Remodel Lipid Metabolism: Strategies Targeting Transcription FactorsHossam El-basiounyNessuna valutazione finora
- 5G Radiation and The COVID-19 Pandemic: Coincidence or Causal RelationshipDocumento3 pagine5G Radiation and The COVID-19 Pandemic: Coincidence or Causal RelationshipMartin L. Pall100% (10)
- The Clinical Potential of Senolytic DrugsDocumento5 pagineThe Clinical Potential of Senolytic DrugsAmadon FaulNessuna valutazione finora
- Ginger: A Functional Herb: January 2013Documento30 pagineGinger: A Functional Herb: January 2013marutheshNessuna valutazione finora
- Mechanisms of Cell InjuryDocumento21 pagineMechanisms of Cell Injuryبراءة أحمد السلاماتNessuna valutazione finora
- Cochlear Implant Research Updates ITO12Documento242 pagineCochlear Implant Research Updates ITO12Kevin CostainNessuna valutazione finora
- Cancer Homeopathy ReviewDocumento8 pagineCancer Homeopathy ReviewAkashlin DevakumarNessuna valutazione finora
- Kuliah Tumor Biology 2016Documento100 pagineKuliah Tumor Biology 2016izulNessuna valutazione finora
- Perillyl Alcohol (Monoterpene Alcohol), LimoneneDocumento26 paginePerillyl Alcohol (Monoterpene Alcohol), LimoneneDanielNessuna valutazione finora
- Curcumin-Free Turmeric Exhibits Anti-Inflammatory and Anticancer Activities: Identification of Novel Components of TurmericDocumento14 pagineCurcumin-Free Turmeric Exhibits Anti-Inflammatory and Anticancer Activities: Identification of Novel Components of TurmericHoàngTuấnNessuna valutazione finora
- Incucyte Annexin VDocumento6 pagineIncucyte Annexin V李琇琴Nessuna valutazione finora
- Pathology Exam Questions FinalDocumento341 paginePathology Exam Questions FinalTasos ZelevarisNessuna valutazione finora
- KIMEDDocumento8 pagineKIMEDAgung MaulanaNessuna valutazione finora
- Saussurea Lappa Review 2Documento12 pagineSaussurea Lappa Review 2Mulayam Singh YadavNessuna valutazione finora
- Contoh Draft Jurnal 1Documento13 pagineContoh Draft Jurnal 1Palakhi SkmNessuna valutazione finora
- Cells 08 01419Documento35 pagineCells 08 01419dzaky fadhilNessuna valutazione finora
- Chronic Obstructive Pulmonary Disease and The Cardiovascular System Vascular Repair and Regeneration As A Therapeutic TargetDocumento26 pagineChronic Obstructive Pulmonary Disease and The Cardiovascular System Vascular Repair and Regeneration As A Therapeutic TargetAndreea MoalesNessuna valutazione finora
- 19181111Documento104 pagine19181111Jennifer HaasNessuna valutazione finora
- Programmed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciDocumento23 pagineProgrammed Cell Death in Plants: Narcin Palavan-Unsal, Elif-Damla Buyuktuncer and Mehmet Ali TufekciSourav GuptaNessuna valutazione finora
- Abrahamse 2016Documento18 pagineAbrahamse 2016Widia Ade SeptianaNessuna valutazione finora
- DR Haryadi Molecular Basis CancerDocumento59 pagineDR Haryadi Molecular Basis CancerAmirul HadiNessuna valutazione finora
- Really Essential Medical ImmunologyDocumento224 pagineReally Essential Medical Immunology99noname100% (2)
- Human Herpes Virus 5 Complete GenomeDocumento86 pagineHuman Herpes Virus 5 Complete GenomeadrianadamjanovicNessuna valutazione finora
- Pathophysiology Final 1Documento162 paginePathophysiology Final 1Yeshaa Mirani100% (1)
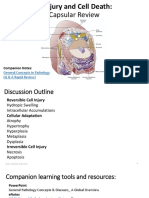
- Cell Injury and Cell DeathDocumento35 pagineCell Injury and Cell DeathMarc Imhotep Cray, M.D.Nessuna valutazione finora
- Anticancer Drugs: Mittwoch, 30. September 2009Documento50 pagineAnticancer Drugs: Mittwoch, 30. September 2009pmfffNessuna valutazione finora
- Alfa Tau Herbal Remedies TestimonialDocumento55 pagineAlfa Tau Herbal Remedies TestimonialЗорица ЋирићNessuna valutazione finora