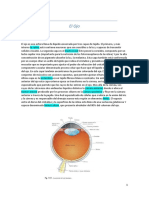
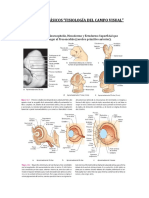
Potrebbero piacerti anche
- Beneficios del Cannabis Medicinal en el GlaucomaDa EverandBeneficios del Cannabis Medicinal en el GlaucomaNessuna valutazione finora
- Sensacion y PercepcionDocumento16 pagineSensacion y PercepcionEstephany Avila ArenasNessuna valutazione finora
- Material Teórico Complementario Visión-2024-CarraDocumento17 pagineMaterial Teórico Complementario Visión-2024-Carrafrancososa63245Nessuna valutazione finora
- Unidad 2 VisiónDocumento8 pagineUnidad 2 VisiónSol AldereteNessuna valutazione finora
- Fisiologia Sensorial de La Via OpticaDocumento45 pagineFisiologia Sensorial de La Via OpticaGabrielaNessuna valutazione finora
- Tarea 3 - Via Visual PDFDocumento30 pagineTarea 3 - Via Visual PDFJaimeSergioVasquezZunigaNessuna valutazione finora
- Neurociencias IIDocumento4 pagineNeurociencias IIAbril ElisNessuna valutazione finora
- 5.3 Fisiología de Entrada de La Luz y Reflejos PupilaresDocumento41 pagine5.3 Fisiología de Entrada de La Luz y Reflejos PupilaresOscar Padilla Navarro100% (1)
- Transduccion VisualDocumento14 pagineTransduccion VisualErika Llanos Mora50% (2)
- Libro Logopedia Trastornos Psicopatologicosdel Lenguaje PDFDocumento152 pagineLibro Logopedia Trastornos Psicopatologicosdel Lenguaje PDFPilarGiralteMonzónNessuna valutazione finora
- F. Práct Sent Esp Ojo 2Documento8 pagineF. Práct Sent Esp Ojo 2Doménica HerediaNessuna valutazione finora
- Biofísica de La Visión AbiudDocumento6 pagineBiofísica de La Visión AbiudAbiud K. AmozurrutiaNessuna valutazione finora
- VisiónDocumento17 pagineVisiónDavidNessuna valutazione finora
- Tema 5Documento8 pagineTema 5LAURA GUERRERO CALDERÓNNessuna valutazione finora
- Trabajo de Fisiología-2Documento5 pagineTrabajo de Fisiología-2Mafer BarrazaNessuna valutazione finora
- Practica #4 MED 411 VisionDocumento4 paginePractica #4 MED 411 VisionFabrizio Mateo Quispe GutiérrezNessuna valutazione finora
- Apuntes Neurofisiologia AutoaprendizajeDocumento18 pagineApuntes Neurofisiologia Autoaprendizajeana lauraNessuna valutazione finora
- Sistema Visual (Resumen)Documento3 pagineSistema Visual (Resumen)Rosendblack100% (1)
- Fisiología de La VisiónDocumento13 pagineFisiología de La VisiónEider Mercado G'amarraNessuna valutazione finora
- Biofísica de La VisiónDocumento9 pagineBiofísica de La VisiónPaola Correa M100% (1)
- Word REtinaDocumento89 pagineWord REtinaCesar Alex Lumbre YuptonNessuna valutazione finora
- NeuroFIsiologia - Vias Visuales CentralesDocumento5 pagineNeuroFIsiologia - Vias Visuales CentralesCRISTOBAL JAVIER LOPEZ ALISTENessuna valutazione finora
- La RetinaDocumento17 pagineLa RetinaEderth Cordero100% (1)
- 2 Sesión Proceso Visual y Percepción VisualDocumento59 pagine2 Sesión Proceso Visual y Percepción VisualChris S. M.Nessuna valutazione finora
- UTA VisiónDocumento62 pagineUTA VisiónAbigail AcurioNessuna valutazione finora
- Tema 4. Psicobiología de La Visión. Versión DefinitivaDocumento10 pagineTema 4. Psicobiología de La Visión. Versión DefinitivaManuel Cañas LucendoNessuna valutazione finora
- Articulo Cientifico de Anatomia Ocular FotorreceptoresDocumento8 pagineArticulo Cientifico de Anatomia Ocular FotorreceptoresLenny ChumoNessuna valutazione finora
- Valoracion PupilarDocumento11 pagineValoracion PupilarNicolasMolanoNessuna valutazione finora
- Vía ÓpticaDocumento14 pagineVía ÓpticaJavier BarretoNessuna valutazione finora
- HolaDocumento4 pagineHolaFernandoNessuna valutazione finora
- Practica Nro 4 Fisiologia de La Visión (24970)Documento6 paginePractica Nro 4 Fisiologia de La Visión (24970)Gabriela GutierrezNessuna valutazione finora
- Vision Diurna.Documento48 pagineVision Diurna.FergatNessuna valutazione finora
- Optica Vision LentesDocumento4 pagineOptica Vision LentesDiego Atoche100% (1)
- Transduccion VisualDocumento7 pagineTransduccion Visualpp0% (1)
- El Ojo HumanoDocumento5 pagineEl Ojo HumanoKarlyHBK100% (1)
- Lab 9-Sentidos de Transducción Física Visión y Equilibrio 2023Documento11 pagineLab 9-Sentidos de Transducción Física Visión y Equilibrio 2023NataliaNessuna valutazione finora
- Extra 3. Sensibilidad VisualDocumento7 pagineExtra 3. Sensibilidad VisualAlbaNessuna valutazione finora
- Sistema Visual El OjoDocumento4 pagineSistema Visual El OjoHeydi AlemánNessuna valutazione finora
- Fundamentos Físicos de La VisiónDocumento14 pagineFundamentos Físicos de La VisiónLaura RamirezNessuna valutazione finora
- Clase 13. Transducción ÓpticaDocumento41 pagineClase 13. Transducción ÓpticaMIGUEL ANGEL PALOMINO CAMPONessuna valutazione finora
- Fisiología de La VisiónDocumento33 pagineFisiología de La VisiónJuan Carlos Monte GonzálezNessuna valutazione finora
- Articulo Cientifico de Los FotorreceptoresDocumento8 pagineArticulo Cientifico de Los FotorreceptoresLenny ChumoNessuna valutazione finora
- Practica 4Documento5 paginePractica 4Drakken gamesNessuna valutazione finora
- Apunte AnatomiaDocumento10 pagineApunte AnatomiaCristian GallardoNessuna valutazione finora
- FOTORRECEPCIÓNDocumento61 pagineFOTORRECEPCIÓNGustavoNessuna valutazione finora
- Potenciales Evocados VisualesDocumento11 paginePotenciales Evocados VisualesCa SalNessuna valutazione finora
- Materia FisioDocumento694 pagineMateria FisioElizabeth HurtadoNessuna valutazione finora
- Carlson 6 VISIÓNDocumento23 pagineCarlson 6 VISIÓNlucasNessuna valutazione finora
- Lobulo OccipitalDocumento93 pagineLobulo OccipitalJairo Nava Briceño100% (4)
- 3er Cuestionario - Fisiologia IDocumento13 pagine3er Cuestionario - Fisiologia IMiguel TejadaNessuna valutazione finora
- Vista ZulyDocumento21 pagineVista ZulyJesus Enrique Rivas CastilloNessuna valutazione finora
- Vias Opticas Proyecto de InvestigacionDocumento5 pagineVias Opticas Proyecto de InvestigacionDenisseNessuna valutazione finora
- Visión - Wikipedia, La Enciclopedia LibreDocumento9 pagineVisión - Wikipedia, La Enciclopedia LibreBeto GomitoNessuna valutazione finora
- Clase de Visión y Audición - Fernando BermejoDocumento103 pagineClase de Visión y Audición - Fernando BermejoPablo GreggioNessuna valutazione finora
- Análisis y Síntesis de Estímulos VisualesDocumento31 pagineAnálisis y Síntesis de Estímulos VisualesPaula GomezNessuna valutazione finora
- Campo VisualDocumento45 pagineCampo VisualGabriela Paz Mengoya Cid100% (1)
- Ojo y Vision SemiologiaDocumento57 pagineOjo y Vision Semiologiaasma_roig100% (1)
- Tema 2 Psicologia FisiologicaDocumento11 pagineTema 2 Psicologia FisiologicaAlvaro Garrido GarciaNessuna valutazione finora
- Anatomía y Fisiología de La VisiónDocumento4 pagineAnatomía y Fisiología de La VisiónMichelle RoblesNessuna valutazione finora
- Celula de MullerDocumento3 pagineCelula de MullerSantkiz StkNessuna valutazione finora
- PLANIFICACION DIARIA YesiDocumento3 paginePLANIFICACION DIARIA Yesiyesenia cruzNessuna valutazione finora
- Academia Amantes Del Conocimiento: Ciclo Repaso San Marcos 2023 - IDocumento11 pagineAcademia Amantes Del Conocimiento: Ciclo Repaso San Marcos 2023 - IDiego Dueñas VelardeNessuna valutazione finora
- Clases de Diagramas para Trabajos EscritosDocumento6 pagineClases de Diagramas para Trabajos Escritosdlugo1973Nessuna valutazione finora
- Rubrica AnaliticaDocumento2 pagineRubrica Analiticaluisa fernanda p. cardonaNessuna valutazione finora
- Cómo Aplicamos La Antropología en La Experiencia de UsuarioDocumento5 pagineCómo Aplicamos La Antropología en La Experiencia de UsuarioKirvis MendezNessuna valutazione finora
- PLAN DE MEJORA FORTIDENT FinalDocumento25 paginePLAN DE MEJORA FORTIDENT FinalARNULFO REYES MUŃOZNessuna valutazione finora
- Qué Son Los Cuerpos RedondosDocumento7 pagineQué Son Los Cuerpos RedondosPame Olivera Reyes0% (1)
- AtmosferaDocumento3 pagineAtmosferaCartru BknNessuna valutazione finora
- "Renta Chingón Tu Airbnb o Uno Ajeno" - Con Estos 5 Sencillos Pasos.Documento6 pagine"Renta Chingón Tu Airbnb o Uno Ajeno" - Con Estos 5 Sencillos Pasos.Lucia del AngelNessuna valutazione finora
- Taller Mecanico Informe CascoDocumento14 pagineTaller Mecanico Informe CascoAriel ChafuelanNessuna valutazione finora
- Marte y El Enigma de La Vida 8Documento2 pagineMarte y El Enigma de La Vida 8Jorge Luis Martinez MazaNessuna valutazione finora
- Empresa Sesion 2Documento23 pagineEmpresa Sesion 2Alexander GutierrezNessuna valutazione finora
- Improving The Safety Performance Nexus A Study On The. TraducidoDocumento23 pagineImproving The Safety Performance Nexus A Study On The. TraducidoCecilia Albujar VeronaNessuna valutazione finora
- Revision ValerianaDocumento53 pagineRevision ValerianaJavier Andres Camo Jimenez100% (1)
- Accion Decisiva Tarea 1 A La 10Documento110 pagineAccion Decisiva Tarea 1 A La 10RAFAEL ALFONSO HERNANDEZ GARCIANessuna valutazione finora
- Psicología Del Consumidor ENTREGADocumento6 paginePsicología Del Consumidor ENTREGAClaudia Patricia Arregoces RealNessuna valutazione finora
- Mexicanos en Las OlimpiadasDocumento23 pagineMexicanos en Las Olimpiadaselvistecnology8Nessuna valutazione finora
- Informe de Laboratorio Ley Del PénduloDocumento8 pagineInforme de Laboratorio Ley Del PénduloIrani Franco ZabaletaNessuna valutazione finora
- El Siga Como Una Mejora de Control en Los Procedimientos Administrativos y Patrimoniales en La Municipalidad Distrital de Pimentel 2021Documento25 pagineEl Siga Como Una Mejora de Control en Los Procedimientos Administrativos y Patrimoniales en La Municipalidad Distrital de Pimentel 2021Laurita GomezNessuna valutazione finora
- Cuestionario de Matematica 3ro BachilleratoDocumento4 pagineCuestionario de Matematica 3ro Bachilleratowilmupon096867100% (2)
- Formato RadicadoDocumento9 pagineFormato RadicadoEdimer CañonNessuna valutazione finora
- Proyecto ProductivoDocumento60 pagineProyecto ProductivoZairha Daniela Peña BalantaNessuna valutazione finora
- Intranet ExtranetDocumento2 pagineIntranet Extranetcarlos100% (1)
- Mantenimiento de Líneas de Transmisión de EnergíaDocumento6 pagineMantenimiento de Líneas de Transmisión de EnergíaEduardo Gamboa Marin100% (2)
- u3EJERCICIO PRACTICO 4Documento27 pagineu3EJERCICIO PRACTICO 4Vany LozanoNessuna valutazione finora
- Maribel y La Extraña FamiliaDocumento8 pagineMaribel y La Extraña FamiliaPaloma GomisNessuna valutazione finora
- Factor VII ActivadoDocumento3 pagineFactor VII ActivadoJessica Tania Trucios RiosNessuna valutazione finora
- 3.riohs AgostoDocumento98 pagine3.riohs AgostoGiselle Alejandra Alfaro CarcajalNessuna valutazione finora
- Politica de CTI 2017 2030-ParaguayDocumento34 paginePolitica de CTI 2017 2030-Paraguayidelin.molinasNessuna valutazione finora
- Eliecer Lema Tarea5 Plan de NegociosDocumento6 pagineEliecer Lema Tarea5 Plan de NegocioseliecerNessuna valutazione finora