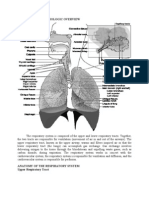
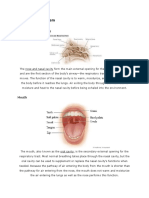
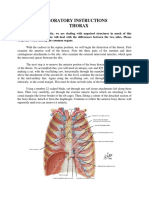
Potrebbero piacerti anche
- Respiratory and Circulatory SystemDocumento10 pagineRespiratory and Circulatory Systemainabukhari20Nessuna valutazione finora
- Plant and Animal Organ System and Their FunctionDocumento21 paginePlant and Animal Organ System and Their FunctionJovon GuyoNessuna valutazione finora
- Anatomy and PhysiologyDocumento5 pagineAnatomy and PhysiologymalindaNessuna valutazione finora
- Concept of Disease 1.1 Anatomy and PhysiologyDocumento15 pagineConcept of Disease 1.1 Anatomy and PhysiologyAulia SandraNessuna valutazione finora
- Respiratory Anatomy Physiology and Dse DefinitionDocumento4 pagineRespiratory Anatomy Physiology and Dse Definitionmiss RN100% (2)
- CH 11 Gas Exchange in HumansDocumento4 pagineCH 11 Gas Exchange in HumansPranitha RaviNessuna valutazione finora
- Anatomy and Physiology Power PointDocumento8 pagineAnatomy and Physiology Power PointJoanna Diongson GonzalesNessuna valutazione finora
- The Vital Respiratory System: Structure and Functions in 37 CharactersDocumento18 pagineThe Vital Respiratory System: Structure and Functions in 37 CharactersShinigami AmnerrorNessuna valutazione finora
- Introduction To Acute Respiratory FailureDocumento6 pagineIntroduction To Acute Respiratory FailureErieca Barsabal PamittanNessuna valutazione finora
- Anatomy and Physiology of The Respiratory SystemDocumento5 pagineAnatomy and Physiology of The Respiratory SystemxoxosvwNessuna valutazione finora
- Breathing and Exchange of GasesDocumento17 pagineBreathing and Exchange of GasesArjun ChristopherNessuna valutazione finora
- The Respiratory SystemDocumento8 pagineThe Respiratory SystemDavid DanielNessuna valutazione finora
- ANATOMY AND PHYSIOLOGY OF THE RESPIRATORY SYSTEMDocumento6 pagineANATOMY AND PHYSIOLOGY OF THE RESPIRATORY SYSTEMCarlos Alfonso Borromeo AmoresNessuna valutazione finora
- GAS EXCHANGE AND THE RESPIRATORY SYSTEMDocumento7 pagineGAS EXCHANGE AND THE RESPIRATORY SYSTEMChenul WickramasingheNessuna valutazione finora
- Anatomy Physiology - MelaneeDocumento5 pagineAnatomy Physiology - MelaneeNikkoBacasonNessuna valutazione finora
- Respiratory System Functions in 5 StepsDocumento16 pagineRespiratory System Functions in 5 Stepshellofrom theothersideNessuna valutazione finora
- Indah Yulianti - 012011041 - Kep A1'20 - Resume Respiratory SystemDocumento6 pagineIndah Yulianti - 012011041 - Kep A1'20 - Resume Respiratory SystemIndah YuliantiNessuna valutazione finora
- Chapter II - Respiratory SystemDocumento3 pagineChapter II - Respiratory SystemIndranil SinhaNessuna valutazione finora
- Respiratory SystemDocumento10 pagineRespiratory Systemapi-305592242Nessuna valutazione finora
- Anatomy and Physiology: Upper Respiratory TractDocumento3 pagineAnatomy and Physiology: Upper Respiratory TractAltea MartirezNessuna valutazione finora
- Anatomy and PhysiologyDocumento11 pagineAnatomy and Physiologyjawn09euclidNessuna valutazione finora
- Breathing Process PDFDocumento3 pagineBreathing Process PDFDianeNessuna valutazione finora
- Respiratory System OverviewDocumento3 pagineRespiratory System Overviewๆรำำทแร หบหฝำมฟNessuna valutazione finora
- Science Senior 2Documento4 pagineScience Senior 2ajichristianbaniadamNessuna valutazione finora
- Group2 (Respiration)Documento12 pagineGroup2 (Respiration)dinaNessuna valutazione finora
- The Respiratory SystemDocumento3 pagineThe Respiratory Systempaulo_camuaNessuna valutazione finora
- Respiratory SystemDocumento31 pagineRespiratory SystemMark Jhosua Austria GalinatoNessuna valutazione finora
- Core Concepts in Anatomy and PhysiologyDocumento50 pagineCore Concepts in Anatomy and PhysiologyMohamed Abdullah KhojaliNessuna valutazione finora
- Respiratory System - Anatomy and PhysiologyDocumento6 pagineRespiratory System - Anatomy and PhysiologyJay Crishnan Morales CajandingNessuna valutazione finora
- OxygenationDocumento20 pagineOxygenationKhie-An OcampoNessuna valutazione finora
- Gas Exchange NewDocumento30 pagineGas Exchange Newmariaamkhaann22Nessuna valutazione finora
- Respiratory System: Anatomy of The NoseDocumento6 pagineRespiratory System: Anatomy of The NoseAlmira ColumbanoNessuna valutazione finora
- Anatomic and Physiologic OverviewDocumento8 pagineAnatomic and Physiologic OverviewJoseph King MacaranasNessuna valutazione finora
- Grade 9 Lesson1Documento60 pagineGrade 9 Lesson1Julius LitaNessuna valutazione finora
- RESPIRATORY SYSTEMDocumento3 pagineRESPIRATORY SYSTEMjoimaciascorreaNessuna valutazione finora
- Respiratory System: Nose and Nasal CavityDocumento8 pagineRespiratory System: Nose and Nasal CavityJasper AlbuferaNessuna valutazione finora
- The Structure of The Respiratory SystemDocumento3 pagineThe Structure of The Respiratory SystemІрина КозакNessuna valutazione finora
- The Cells of The Human Body Require A Constant Stream of Oxygen To Stay AliveDocumento14 pagineThe Cells of The Human Body Require A Constant Stream of Oxygen To Stay AlivePeperoniiNessuna valutazione finora
- Iv. Anatomy and Physiology of The Human Respiratory SystemDocumento5 pagineIv. Anatomy and Physiology of The Human Respiratory SystemJenny Vi CodenieraNessuna valutazione finora
- Parts and Functions of Respiratory SystemDocumento18 pagineParts and Functions of Respiratory Systemjosephabram051590Nessuna valutazione finora
- Anatomy and PhysiologyDocumento1 paginaAnatomy and PhysiologyDian Rose Sebanes MamintaNessuna valutazione finora
- Essential Functions of the Respiratory SystemDocumento5 pagineEssential Functions of the Respiratory SystemEjie Boy IsagaNessuna valutazione finora
- Parts and Functions of Respiratory System.Documento3 pagineParts and Functions of Respiratory System.Alvin Patrick Colobong AsisNessuna valutazione finora
- Respiratory System OverviewDocumento19 pagineRespiratory System OverviewTsume ShoutaroNessuna valutazione finora
- Anatomy and Physiology: PHYSIOLOGY-the Branch of Biology That Deals With The Internal Workings ofDocumento4 pagineAnatomy and Physiology: PHYSIOLOGY-the Branch of Biology That Deals With The Internal Workings ofshonievin100% (2)
- Respiratory System: Anatomy of Respiratory Organs and Their FunctionsDocumento6 pagineRespiratory System: Anatomy of Respiratory Organs and Their FunctionsRishabh SharmaNessuna valutazione finora
- The human respiratory system: how breathing worksDocumento8 pagineThe human respiratory system: how breathing worksChilla Adis ViolitaNessuna valutazione finora
- Anatomy and Physiology Assignment 3Documento6 pagineAnatomy and Physiology Assignment 3api-238869728Nessuna valutazione finora
- Anatomy and Physiology of The Respiratory SystemDocumento3 pagineAnatomy and Physiology of The Respiratory SystemLyka100% (1)
- Study Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!Documento17 pagineStudy Materials: Vedantu Innovations Pvt. Ltd. Score High With A Personal Teacher, Learn LIVE Online!Saraswati PNessuna valutazione finora
- Respiratory System VocabularyDocumento9 pagineRespiratory System VocabularyAgustín Bravo ArreyesNessuna valutazione finora
- Research PaperDocumento14 pagineResearch PaperAlicia ValverdeNessuna valutazione finora
- ReproductiveDocumento5 pagineReproductiveJason Vinluan CarinanNessuna valutazione finora
- A & P RespiratoryDocumento18 pagineA & P Respiratoryamirul haakimNessuna valutazione finora
- Respiratory System ExplainedDocumento11 pagineRespiratory System ExplainedMark Dave VillasencioNessuna valutazione finora
- Respiratory SystemDocumento54 pagineRespiratory SystemALYZZA JANE BUNANessuna valutazione finora
- Oxygenation: Anatomy and PhysiologyDocumento2 pagineOxygenation: Anatomy and PhysiologyMark Elen100% (1)
- Parts & Function of Respiratory SystemDocumento4 pagineParts & Function of Respiratory SystemLucille Ballares83% (6)
- Notes 1Documento12 pagineNotes 1Muhammad TaimoorNessuna valutazione finora
- How Do Humans Breathe? Science Book Age 8 | Children's Biology BooksDa EverandHow Do Humans Breathe? Science Book Age 8 | Children's Biology BooksNessuna valutazione finora
- Respiratory EmergenciesDocumento30 pagineRespiratory EmergenciesNovriefta NugrahaNessuna valutazione finora
- Fraktur Iga PosteriorDocumento8 pagineFraktur Iga PosteriorJulia Kasab 1Nessuna valutazione finora
- Human Anatomy Physiology Laboratory Manual Fetal Pig Version 12th Edition Marieb Solution ManualDocumento5 pagineHuman Anatomy Physiology Laboratory Manual Fetal Pig Version 12th Edition Marieb Solution Manualsritrader89% (9)
- MerKaBa Meditation - Drunvalo MelchizedekDocumento8 pagineMerKaBa Meditation - Drunvalo MelchizedekAnjee SugatriNessuna valutazione finora
- Chest Tube Care and Drainage SystemsDocumento3 pagineChest Tube Care and Drainage SystemsDiana CalderonNessuna valutazione finora
- Anatomy Physiology Disease 3rd Colbert Test BankDocumento10 pagineAnatomy Physiology Disease 3rd Colbert Test BankfahedqNessuna valutazione finora
- Medical Terminlogy Body Planes SectionsDocumento75 pagineMedical Terminlogy Body Planes SectionsSTEM-G.04 Kiarrah Katrina BotinNessuna valutazione finora
- Short Cases in MedicineDocumento30 pagineShort Cases in MedicineselamuNessuna valutazione finora
- Lesson Five 4 - 5837179529600176303Documento9 pagineLesson Five 4 - 5837179529600176303julien KoudouNessuna valutazione finora
- Full Body Reflexology Chart Template Free PDF Format PDFDocumento37 pagineFull Body Reflexology Chart Template Free PDF Format PDFShadrina Al Shafama100% (3)
- Trauma patient management stepsDocumento7 pagineTrauma patient management stepsNiceoNessuna valutazione finora
- Handout Advanced Course in Soft Tissue Surgery 2011Documento64 pagineHandout Advanced Course in Soft Tissue Surgery 2011Mihaela AndreiNessuna valutazione finora
- Case Study (Acute Pyelonephritis) FINALDocumento20 pagineCase Study (Acute Pyelonephritis) FINALian-movilla-751780% (10)
- Manual de Ingles de Medicina CompletoDocumento138 pagineManual de Ingles de Medicina CompletoYobelkis Paulino0% (1)
- Modul Anatomi Blok 7Documento27 pagineModul Anatomi Blok 7Dewandaru I A BNessuna valutazione finora
- Chest X-Ray InterpretationDocumento30 pagineChest X-Ray InterpretationNuru99100% (19)
- Part 7 Anterior Thorax WallDocumento2 paginePart 7 Anterior Thorax Walldragoonm9221Nessuna valutazione finora
- Rapid Medical AssessmentDocumento2 pagineRapid Medical AssessmentJan MitchelleNessuna valutazione finora
- Artefacts: DR. T. T. K. ReddyDocumento38 pagineArtefacts: DR. T. T. K. ReddyBolisetty HemanthNessuna valutazione finora
- Homeopathy BRYONIA ALBA - COMPLETE Information About BRYONIA ALBA - Samuel HahnemannDocumento36 pagineHomeopathy BRYONIA ALBA - COMPLETE Information About BRYONIA ALBA - Samuel HahnemannSorin AilincaiNessuna valutazione finora
- Wilkins Clinical Assessment in Respiratory Care 7Th Edition Heuer Test Bank Full Chapter PDFDocumento31 pagineWilkins Clinical Assessment in Respiratory Care 7Th Edition Heuer Test Bank Full Chapter PDFmichellesmith25081983ebo100% (5)
- Impaired Gas ExchangeDocumento3 pagineImpaired Gas ExchangePaul VincentNessuna valutazione finora
- 2020thoraxlungs Neckheart VesselsDocumento171 pagine2020thoraxlungs Neckheart VesselsFaith madayagNessuna valutazione finora
- PERFORMANCE-EVALUATION-CHECKLIST-HEADDocumento4 paginePERFORMANCE-EVALUATION-CHECKLIST-HEADHaydi Pineda MedinaNessuna valutazione finora
- Anaesthetic Management of Lumber PivdDocumento21 pagineAnaesthetic Management of Lumber PivdSadasiv PandaNessuna valutazione finora
- MODULE 4-Do's and Don'Ts in Physical Activities and Exercise Proper Breathing and Warm-Up - StretchingDocumento65 pagineMODULE 4-Do's and Don'Ts in Physical Activities and Exercise Proper Breathing and Warm-Up - StretchingAljeya EvanchezNessuna valutazione finora
- Explosion and Damage Assessment Computer Simulation Using HExDAM PDFDocumento6 pagineExplosion and Damage Assessment Computer Simulation Using HExDAM PDFYonish SisoNessuna valutazione finora
- Side Effects of Radiation TherapyDocumento7 pagineSide Effects of Radiation TherapySerrano Ivan JasperNessuna valutazione finora
- FCA-ppt DefenseDocumento72 pagineFCA-ppt DefenseclarNessuna valutazione finora
- Tamil For MedicosDocumento32 pagineTamil For MedicosKaveri Somasundaram100% (1)