Potrebbero piacerti anche
- The Concise Human Body Book An Illustrated Guide To Its Structure, Function, and Disorders PDFDocumento322 pagineThe Concise Human Body Book An Illustrated Guide To Its Structure, Function, and Disorders PDFBriyith Triana Rondon100% (4)
- Newport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALDocumento8 pagineNewport, D.J. and Nemeroff, C.B. Neurobiology of Posttraumatic Stress Disorder. Cognitive Neuroscience JAARTALg10564433Nessuna valutazione finora
- (Eisenberg@@) (2000) (NA) (X) Protein Function PDFDocumento4 pagine(Eisenberg@@) (2000) (NA) (X) Protein Function PDFbyominNessuna valutazione finora
- Art7 PDFDocumento11 pagineArt7 PDFKatthy AndrangoNessuna valutazione finora
- Research Paper On Protein BindingDocumento7 pagineResearch Paper On Protein Bindingnadevufatuz2100% (1)
- Astrina Spectro Masas 2001Documento6 pagineAstrina Spectro Masas 2001Nacido para BendcirNessuna valutazione finora
- PIC-Seq Dissecting Cellular Crosstalk by Sequencing Physically Interacting CellsDocumento16 paginePIC-Seq Dissecting Cellular Crosstalk by Sequencing Physically Interacting CellsNiv SabathNessuna valutazione finora
- Complejidas Genoma OrganismosDocumento4 pagineComplejidas Genoma OrganismosMamen Casado MuñozNessuna valutazione finora
- The Interplay Between Disordered Regions in Rnas and Proteins Modulates Interactions Within Stress Granules and Processing BodiesDocumento12 pagineThe Interplay Between Disordered Regions in Rnas and Proteins Modulates Interactions Within Stress Granules and Processing BodiesAngelica Maria Torregroza EspinosaNessuna valutazione finora
- MBSS 2011 Program With AbstractsDocumento12 pagineMBSS 2011 Program With AbstractsphotopidgeNessuna valutazione finora
- Detecting Protein-Protein Interactions by Far Western BlottingDocumento7 pagineDetecting Protein-Protein Interactions by Far Western BlottingJinsheng ZhuNessuna valutazione finora
- Proteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)Documento306 pagineProteins and Cell Regulation Vol 03 - RHO Family GTPases, 1E (2005)DiahaNessuna valutazione finora
- Toylife: A Computational Framework To Study The Multi-Level Organisation of The Genotype-Phenotype MapDocumento10 pagineToylife: A Computational Framework To Study The Multi-Level Organisation of The Genotype-Phenotype MapdonlockwoodNessuna valutazione finora
- Modification of Drosophila p53 by SUMO Modulates Its Transactivation and Pro-Apoptotic FunctionsDocumento9 pagineModification of Drosophila p53 by SUMO Modulates Its Transactivation and Pro-Apoptotic FunctionsGustavo FelpeNessuna valutazione finora
- Protein Interaction Maps Using Yeast Two-Hybrid Assay: Shayantani Mukherjee, Sampali Bal and Partha SahaDocumento7 pagineProtein Interaction Maps Using Yeast Two-Hybrid Assay: Shayantani Mukherjee, Sampali Bal and Partha SahamriduNessuna valutazione finora
- Chaperon AsDocumento6 pagineChaperon AsVale GerónimoNessuna valutazione finora
- Properties and Functions ofDocumento18 pagineProperties and Functions ofNivetha NagarajanNessuna valutazione finora
- The Hepatitis B Virus Pres1 Domain Hijacks Host Trafficking Proteins by Motif MimicryDocumento10 pagineThe Hepatitis B Virus Pres1 Domain Hijacks Host Trafficking Proteins by Motif MimicrySumit MukherjeeNessuna valutazione finora
- Mini JoseDocumento14 pagineMini JoseAnita BatistaNessuna valutazione finora
- Pi Is 0006349518321933Documento1 paginaPi Is 0006349518321933ASP ValenciaNessuna valutazione finora
- TMP 3Documento3 pagineTMP 3FrontiersNessuna valutazione finora
- MED 111 FA-2 Answers 23-24Documento12 pagineMED 111 FA-2 Answers 23-24ozerbilge24Nessuna valutazione finora
- Arabidopsis Thaliana: Designing A Computational System To Predict Protein-Protein Interactions inDocumento4 pagineArabidopsis Thaliana: Designing A Computational System To Predict Protein-Protein Interactions inbikashNessuna valutazione finora
- 1 s2.0 S0006349518316114 MainDocumento1 pagina1 s2.0 S0006349518316114 MainDiego TulcanNessuna valutazione finora
- The C. Elegans F-box/WD-repeat Protein LIN-23 Functions To Limit Cell Division During DevelopmentDocumento12 pagineThe C. Elegans F-box/WD-repeat Protein LIN-23 Functions To Limit Cell Division During DevelopmentStrahil NikolovNessuna valutazione finora
- Papers Abstract To ReadDocumento2 paginePapers Abstract To ReadSapan MandloiNessuna valutazione finora
- Genes and ProteinsDocumento3 pagineGenes and ProteinsVina Palce-BobilaNessuna valutazione finora
- 10.1038@s41573 020 0063 yDocumento13 pagine10.1038@s41573 020 0063 ySpoti PremiNessuna valutazione finora
- Transcriptome Analysis Reveals Dysregulation of Innate Immune Response Genes and Neuronal Activity Dependent Genes in AutismDocumento8 pagineTranscriptome Analysis Reveals Dysregulation of Innate Immune Response Genes and Neuronal Activity Dependent Genes in AutismKarim ElghachtoulNessuna valutazione finora
- Robust Spatial Sensing of Mating Pheromone Gradients by Yeast CellsDocumento11 pagineRobust Spatial Sensing of Mating Pheromone Gradients by Yeast CellsTau-Mu YiNessuna valutazione finora
- Viale 2014Documento9 pagineViale 2014Maria Laura MonNessuna valutazione finora
- AdhesionDocumento4 pagineAdhesionnuman88Nessuna valutazione finora
- Immunoglobulin Deposition On Biomolecule Corona Determines Complement Opsonization Efficiency of Preclinical and Clinical NanoparticlesDocumento14 pagineImmunoglobulin Deposition On Biomolecule Corona Determines Complement Opsonization Efficiency of Preclinical and Clinical NanoparticlesRyad HBNessuna valutazione finora
- Host-Pathogen Interactions: The Seduction of Molecular Cross TalkDocumento7 pagineHost-Pathogen Interactions: The Seduction of Molecular Cross TalkPoppy Siska IsabellaNessuna valutazione finora
- 1059 FullDocumento19 pagine1059 FullCARLOS FRANCISCO MANTILLA MONTALVONessuna valutazione finora
- Chapter 11 Gene ExpressionDocumento30 pagineChapter 11 Gene ExpressionJonathan WyattNessuna valutazione finora
- Thesis On Gene Expression AnalysisDocumento5 pagineThesis On Gene Expression Analysisbrittanyjonescolumbia100% (1)
- A Biochemical Network Modeling of A Whole CellDocumento14 pagineA Biochemical Network Modeling of A Whole CellabdurrehmanNessuna valutazione finora
- Assembly of Protein Complexes Restricts DiffusionDocumento15 pagineAssembly of Protein Complexes Restricts DiffusionRifqi Fathul ArroisiNessuna valutazione finora
- 1 s2.0 S0006349513015762 Main PDFDocumento1 pagina1 s2.0 S0006349513015762 Main PDFDiego TulcanNessuna valutazione finora
- Micro Algae The Syncronized EukaryotesDocumento2 pagineMicro Algae The Syncronized EukaryotesWillyYeeNessuna valutazione finora
- BMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in MiceDocumento14 pagineBMC Biotechnology: Dynamic in Vivo Imaging and Cell Tracking Using A Histone Fluorescent Protein Fusion in Micefather45Nessuna valutazione finora
- Evolution of A Minimal Cell: ArticleDocumento24 pagineEvolution of A Minimal Cell: ArticleJoão VictorNessuna valutazione finora
- Basics of ProteomicsDocumento7 pagineBasics of ProteomicskonndaaiahNessuna valutazione finora
- Viral Regulation of Host Cell Biology by Hijacking of The Nucleolar DNA-damage ResponseDocumento13 pagineViral Regulation of Host Cell Biology by Hijacking of The Nucleolar DNA-damage ResponseDeepa V.DNessuna valutazione finora
- Protein Contact Prediction From Amino Acid Co-Evolution Using Convolutional Networks For Graph-Valued ImagesDocumento9 pagineProtein Contact Prediction From Amino Acid Co-Evolution Using Convolutional Networks For Graph-Valued ImagesThao LeNessuna valutazione finora
- Pro Tea SomeDocumento7 paginePro Tea SomeJuan Garzón LamarqueNessuna valutazione finora
- TMP E289Documento12 pagineTMP E289FrontiersNessuna valutazione finora
- Quantification of Protein Half-Lives in The Budding Yeast ProteomeDocumento6 pagineQuantification of Protein Half-Lives in The Budding Yeast ProteomesecchanNessuna valutazione finora
- Protein Synthesis in CellDocumento7 pagineProtein Synthesis in Cellkvpyqegld100% (2)
- Bergen Dahl 2019Documento12 pagineBergen Dahl 2019Noor ShahizanNessuna valutazione finora
- Cellular Model System To Dissect The Isoform-Selectivity of Akt InhibitorsDocumento14 pagineCellular Model System To Dissect The Isoform-Selectivity of Akt InhibitorsPaing KhantNessuna valutazione finora
- Paper P53Documento18 paginePaper P53LidiaAlejandraRiveraPonceNessuna valutazione finora
- Genes 13 00165 With CoverDocumento12 pagineGenes 13 00165 With CoverMaria Garcia FernandezNessuna valutazione finora
- Three Dimensional Genome Architecture Players and MechanismsDocumento13 pagineThree Dimensional Genome Architecture Players and MechanismsNatalio Vazquez CruzNessuna valutazione finora
- Walking The Interactome For Prioritization of Candidate Disease GenesDocumento10 pagineWalking The Interactome For Prioritization of Candidate Disease GenesLiza MarcelNessuna valutazione finora
- Computational Proteomics: High-Throughput Analysis For Systems BiologyDocumento6 pagineComputational Proteomics: High-Throughput Analysis For Systems BiologyMuhammad TahirNessuna valutazione finora
- Modeling Alzheimer's...Documento35 pagineModeling Alzheimer's...clfp25Nessuna valutazione finora
- Protein Membrane Overlay AssayDocumento4 pagineProtein Membrane Overlay AssayZain BaderNessuna valutazione finora
- Topical Guidebook For GCE O Level Biology 3 Part 2Da EverandTopical Guidebook For GCE O Level Biology 3 Part 2Valutazione: 5 su 5 stelle5/5 (1)
- Mechanisms of Eukaryotic DNA RecombinationDa EverandMechanisms of Eukaryotic DNA RecombinationMax E GottesmanNessuna valutazione finora
- 22 Stretches To Improve Your Flexibility For PoleDocumento10 pagine22 Stretches To Improve Your Flexibility For PoleSarah86% (7)
- Resp AssessmentDocumento19 pagineResp AssessmentNURSES- NOOK & CORNERNessuna valutazione finora
- 11 Biology Notes ch05 Morphology of Flowering Plants PDFDocumento7 pagine11 Biology Notes ch05 Morphology of Flowering Plants PDFMeenakshi VenkataramanNessuna valutazione finora
- Taller 8Documento2 pagineTaller 8monchito83hotmailesNessuna valutazione finora
- II. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCDocumento12 pagineII. Blood Vessels Physiology By: Dr. Abdulrahman Aqra MD MSCapi-290338101Nessuna valutazione finora
- Care of Clients With Neurologic Deficit: By: Elmer G. Organia, RNDocumento233 pagineCare of Clients With Neurologic Deficit: By: Elmer G. Organia, RNTina TalmadgeNessuna valutazione finora
- Implikasi Pasien Dengan Penyakit Paru Pada PenerbanganDocumento47 pagineImplikasi Pasien Dengan Penyakit Paru Pada PenerbanganFebi Chyanx ElsanabiprilNessuna valutazione finora
- U.S. Army Survival Manual FM 21-76 (June 1992)Documento572 pagineU.S. Army Survival Manual FM 21-76 (June 1992)explorer_ebook100% (1)
- Cardiology & Ekgs: Archer Nclex ReviewDocumento59 pagineCardiology & Ekgs: Archer Nclex Reviewdaphne farleyNessuna valutazione finora
- MIO MediumDocumento2 pagineMIO MediumIntan Farida YasminNessuna valutazione finora
- FluidTx MaintenanceDocumento22 pagineFluidTx MaintenanceJonathan IngramNessuna valutazione finora
- Fudamentals of Biomechanics and Biomechanics of Levelling and Aligning (Includes Biomechanics of Molar Control)Documento326 pagineFudamentals of Biomechanics and Biomechanics of Levelling and Aligning (Includes Biomechanics of Molar Control)Margareth Brito100% (4)
- Anatomy AIIMSDocumento18 pagineAnatomy AIIMSvkNessuna valutazione finora
- Bloody Tears Following Lefort 1Documento3 pagineBloody Tears Following Lefort 1paulwhite1234Nessuna valutazione finora
- Edexcel GCSE Science Topic B1.1 Classification, Variation and Inheritance Test 13 - 14Documento10 pagineEdexcel GCSE Science Topic B1.1 Classification, Variation and Inheritance Test 13 - 14Paul BurgessNessuna valutazione finora
- Uropati ObstruktifDocumento22 pagineUropati ObstruktifEvan Permana100% (1)
- Cell Size Shape and SAV Surface-Area VolumeDocumento8 pagineCell Size Shape and SAV Surface-Area VolumeAylin Gutiérrez JuárezNessuna valutazione finora
- B1 Quiz 12Documento2 pagineB1 Quiz 12giulianotboNessuna valutazione finora
- EMG Methods For Evaluating Muscle and Nerve FunctionDocumento546 pagineEMG Methods For Evaluating Muscle and Nerve Function4569077100% (1)
- Mitochondrial Cytopathies in Children and AdultsDocumento28 pagineMitochondrial Cytopathies in Children and AdultsmmaitehmdNessuna valutazione finora
- As Biology With Stafford Unit 3 Workbook Answers PDFDocumento21 pagineAs Biology With Stafford Unit 3 Workbook Answers PDFFatma ZorluNessuna valutazione finora
- The Peripheral Nervous System Branches Throughout The Body Worksheet1Documento2 pagineThe Peripheral Nervous System Branches Throughout The Body Worksheet1Shekeyva SmalleyNessuna valutazione finora
- Bronchiectasis Physiotherapy Assessment Form 2Documento4 pagineBronchiectasis Physiotherapy Assessment Form 2jefferyNessuna valutazione finora
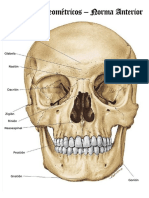
- Puntos Craneometricos CompressDocumento13 paginePuntos Craneometricos CompressLaura PNessuna valutazione finora
- Effect of Plant Growth Enhancers On Growth and Flowering of Tuberose Cv. Prajwal PDFDocumento4 pagineEffect of Plant Growth Enhancers On Growth and Flowering of Tuberose Cv. Prajwal PDFSupal DesaiNessuna valutazione finora
- Format Ketikan LabDocumento9 pagineFormat Ketikan Labapril babyNessuna valutazione finora
- Drug StudyDocumento3 pagineDrug StudyStephannie MirandaNessuna valutazione finora
- Body Systems Portfolio - Tommy JDocumento8 pagineBody Systems Portfolio - Tommy Japi-554072790Nessuna valutazione finora