Potrebbero piacerti anche
- Enseñar evolución y genética para la alfabetización científicaDa EverandEnseñar evolución y genética para la alfabetización científicaNessuna valutazione finora
- BiologÍa Evolutiva Del DesarrolloDocumento9 pagineBiologÍa Evolutiva Del Desarrollomarilynday12Nessuna valutazione finora
- Que Es La Epigenetica What Is EpigenetiDocumento25 pagineQue Es La Epigenetica What Is EpigenetiEmiliano Sevilleja MartinezNessuna valutazione finora
- El Problema Del Aprendizaje en Psicología 2014Documento26 pagineEl Problema Del Aprendizaje en Psicología 2014hercho_zaldaNessuna valutazione finora
- Dialnet GeneYMutacion 62135Documento20 pagineDialnet GeneYMutacion 62135LenardR.Apaza100% (1)
- Evolución y desarrollo biológicoDocumento27 pagineEvolución y desarrollo biológicoJosep Ramon FrasquetNessuna valutazione finora
- Analisis Critico A La LecturaDocumento3 pagineAnalisis Critico A La LecturaMartin CrisantoNessuna valutazione finora
- Era Genetica, Nacimiento de Una Supremacia FinalDocumento22 pagineEra Genetica, Nacimiento de Una Supremacia FinalANGELICA NOGUESNessuna valutazione finora
- GenéticaDocumento18 pagineGenéticaLuisEnriqueVillanuevaSolisNessuna valutazione finora
- Biología EvolutivaDocumento5 pagineBiología EvolutivaiOSvaldo 7Nessuna valutazione finora
- Neodarwinismo o Teoria SinteticaDocumento4 pagineNeodarwinismo o Teoria Sinteticajosue P.M.Nessuna valutazione finora
- Sinopsis de Evolución en Cuatro DimensionesDocumento23 pagineSinopsis de Evolución en Cuatro DimensionesSamuel SantibañezNessuna valutazione finora
- Evolución y Sus ClasificacionesDocumento6 pagineEvolución y Sus Clasificacionesjessfey ggNessuna valutazione finora
- Orígenes y definiciones de la epigenéticaDocumento4 pagineOrígenes y definiciones de la epigenéticaarcimarieNessuna valutazione finora
- Evidencias de La Evolucion - 6°añoDocumento5 pagineEvidencias de La Evolucion - 6°añoDaríoNessuna valutazione finora
- Evo Devo Hacia Un Nuevo Paradigma en Biolog A EvolutivaDocumento3 pagineEvo Devo Hacia Un Nuevo Paradigma en Biolog A EvolutivamagiqoNessuna valutazione finora
- Evolución - Lectura 2Documento10 pagineEvolución - Lectura 2Alejandro SaavedraNessuna valutazione finora
- Evo-Devo: Hacia un nuevo paradigma en Biología EvolutivaDocumento4 pagineEvo-Devo: Hacia un nuevo paradigma en Biología EvolutivaMer OyhNessuna valutazione finora
- El Proyecto Genoma HumanoDocumento1 paginaEl Proyecto Genoma Humanoodalis monroyNessuna valutazione finora
- De Asúa-EvoluciónDocumento22 pagineDe Asúa-EvoluciónMarian MagNessuna valutazione finora
- Introducción A La GenéticaDocumento14 pagineIntroducción A La Genéticaadelina hernandezNessuna valutazione finora
- 17.dialnet InterrogantesPlanteadosPorLaManipulacionGeneticaYE 909351 PDFDocumento24 pagine17.dialnet InterrogantesPlanteadosPorLaManipulacionGeneticaYE 909351 PDFChikorita ChikoritaNessuna valutazione finora
- TEORIAS Y MECANISMOS DE LA EVOLUCION (Recuperado Automáticamente)Documento17 pagineTEORIAS Y MECANISMOS DE LA EVOLUCION (Recuperado Automáticamente)MARTHA LILIANA MEDINA PADILLANessuna valutazione finora
- Filosofia de La BiologiaDocumento15 pagineFilosofia de La BiologiaJaime Andres Vasquez MarchantNessuna valutazione finora
- Importancia de La Teoría de La Evolución en MedicinaDocumento6 pagineImportancia de La Teoría de La Evolución en MedicinaDaniel Eduardo Morán HernándezNessuna valutazione finora
- Biologia Integrativa - AlonzoDocumento10 pagineBiologia Integrativa - AlonzoAdrian PigaleNessuna valutazione finora
- EpigenesisDocumento12 pagineEpigenesistanichusNessuna valutazione finora
- La EvoluciónDocumento4 pagineLa EvoluciónVerónica MartinezNessuna valutazione finora
- La evolución del concepto de gen desde la biología a la ideologíaDocumento14 pagineLa evolución del concepto de gen desde la biología a la ideologíaLucas RoldánNessuna valutazione finora
- Trastornos Mentales y EpigenéticaDocumento29 pagineTrastornos Mentales y EpigenéticaChuy CastañedaNessuna valutazione finora
- Los Hitos Del Conocimiento BiológicoDocumento5 pagineLos Hitos Del Conocimiento BiológicoJuan Manuel Castro Pérez100% (1)
- Mayr Especies Animales y Evolución Cap 1Documento23 pagineMayr Especies Animales y Evolución Cap 1Gonzalo Valenzuela HurtadoNessuna valutazione finora
- Dictado Genetica-Usc PDFDocumento168 pagineDictado Genetica-Usc PDFAnabell Mercado100% (1)
- Investigación DocumentalDocumento19 pagineInvestigación DocumentalLuisNessuna valutazione finora
- James Dewey WatsonDocumento3 pagineJames Dewey WatsonMaría Alicia Pariona HernándezNessuna valutazione finora
- History of Genetics and GenomicsDocumento22 pagineHistory of Genetics and GenomicsNube UrgilezNessuna valutazione finora
- James Dewey Watson, biólogo estadounidenseDocumento3 pagineJames Dewey Watson, biólogo estadounidenseMariali ParionaNessuna valutazione finora
- Ensayo GeneticaDocumento4 pagineEnsayo GeneticaLorena CastilloNessuna valutazione finora
- La GenéticaDocumento12 pagineLa GenéticaWylerNessuna valutazione finora
- 75EUC Genes NicolasJouveDocumento9 pagine75EUC Genes NicolasJouveCatherine DeclercqNessuna valutazione finora
- Desarrollo Historico de La Genetica de PoblacionesDocumento3 pagineDesarrollo Historico de La Genetica de Poblacionesenith johanaNessuna valutazione finora
- M16S1AI1Documento5 pagineM16S1AI1Brian Acevedo50% (2)
- M16S1AI1Documento5 pagineM16S1AI1Brian AcevedoNessuna valutazione finora
- M16S1AI1Documento5 pagineM16S1AI1Brian AcevedoNessuna valutazione finora
- Investigadores George Wells Beadle y Edward Lawrie TatumDocumento7 pagineInvestigadores George Wells Beadle y Edward Lawrie TatumKaren ApazaNessuna valutazione finora
- Entrevista A Massimo PigliucciDocumento3 pagineEntrevista A Massimo PigliucciCarlos DominguezNessuna valutazione finora
- CONCEPTOS BASICOS DE GENETICA 23 2Documento57 pagineCONCEPTOS BASICOS DE GENETICA 23 2Natalia MamaniNessuna valutazione finora
- Teoría de La Evolución Aristoteles, Darwin, LamarckDocumento18 pagineTeoría de La Evolución Aristoteles, Darwin, LamarckKarla Osorio67% (3)
- GENETICADocumento11 pagineGENETICALupitha MendozaNessuna valutazione finora
- Paradigm AsDocumento2 pagineParadigm AsHobi. 140 (fridajikook)Nessuna valutazione finora
- T. 1 Introducción A La GenéticaDocumento5 pagineT. 1 Introducción A La GenéticaYearlin Rodriguez FortunaNessuna valutazione finora
- La Clonacion y La Ingenieria GeneticaDocumento5 pagineLa Clonacion y La Ingenieria GeneticaManuel Zometa GuevaraNessuna valutazione finora
- Definición y Concepto de BiologíaDocumento19 pagineDefinición y Concepto de BiologíaAna SandovalNessuna valutazione finora
- Ética ProfesionalDocumento15 pagineÉtica ProfesionalMony FloresNessuna valutazione finora
- Descubrimiento Del AdnDocumento12 pagineDescubrimiento Del Adnanahi juarezNessuna valutazione finora
- Manipulación GenéticaDocumento3 pagineManipulación GenéticaChristianLópezNessuna valutazione finora
- Biologia Iam G.ocDocumento7 pagineBiologia Iam G.ocGerardo Ortiz CarrilloNessuna valutazione finora
- Explorando los genes: Del big-bang a la nueva BiologíaDa EverandExplorando los genes: Del big-bang a la nueva BiologíaValutazione: 4 su 5 stelle4/5 (7)
- Muy BuenasDocumento11 pagineMuy BuenasAna SbattellaNessuna valutazione finora
- El Indígena en El Imaginario Iconográfico.Documento303 pagineEl Indígena en El Imaginario Iconográfico.Cristian Vargas Paillahueque100% (2)
- Maria Luz EndereDocumento7 pagineMaria Luz EndereAna SbattellaNessuna valutazione finora
- PeriodizaciónDocumento14 paginePeriodizaciónAna SbattellaNessuna valutazione finora
- Programa Sociologia de La S Violencia S Eugenia CozziDocumento12 paginePrograma Sociologia de La S Violencia S Eugenia CozziAna SbattellaNessuna valutazione finora
- VelandiaDocumento18 pagineVelandiaAna SbattellaNessuna valutazione finora
- Soc Poli 13 PDFDocumento2 pagineSoc Poli 13 PDFAna SbattellaNessuna valutazione finora
- Los Orígenes de La Imagen Moderna Del MundoDocumento123 pagineLos Orígenes de La Imagen Moderna Del MundoAna SbattellaNessuna valutazione finora
- Uba Ffyl T 2004 50776 PDFDocumento385 pagineUba Ffyl T 2004 50776 PDFAna SbattellaNessuna valutazione finora
- Gruzinski El Pensamiento Mestizo Cap 3 4 PDFDocumento25 pagineGruzinski El Pensamiento Mestizo Cap 3 4 PDFjsebfagua0% (2)
- HigienismoDocumento2 pagineHigienismoAna SbattellaNessuna valutazione finora
- SV BourgoisDocumento15 pagineSV BourgoisAna SbattellaNessuna valutazione finora
- Efectos GlobalizacionDocumento10 pagineEfectos GlobalizacionlorenceleNessuna valutazione finora
- Propuesta Metodologica para La Identific PDFDocumento145 paginePropuesta Metodologica para La Identific PDFAna SbattellaNessuna valutazione finora
- Peri Rossello 2010Documento13 paginePeri Rossello 2010Ana SbattellaNessuna valutazione finora
- Origen de Las Civilizaciones Antiguas - Jorge E.T Silva Sifuentes PDFDocumento218 pagineOrigen de Las Civilizaciones Antiguas - Jorge E.T Silva Sifuentes PDFJose Andres Zuñiga Cazorla88% (17)
- El Trabajo en La Antiguedad Utiles y Herramientas Serie Guias Didacticas Del Museo Arqueologico Nacional 6Documento49 pagineEl Trabajo en La Antiguedad Utiles y Herramientas Serie Guias Didacticas Del Museo Arqueologico Nacional 6maria ponceNessuna valutazione finora
- Matza Delincuencia y DerivaDocumento30 pagineMatza Delincuencia y DerivaJosela Garcilazo75% (4)
- La Iglesia ColonialDocumento2 pagineLa Iglesia ColonialAna SbattellaNessuna valutazione finora
- Paradigm AsDocumento6 pagineParadigm AsDorthy ColmenaresNessuna valutazione finora
- Yudmila Reyes El VestidoDocumento9 pagineYudmila Reyes El VestidoAna SbattellaNessuna valutazione finora
- Antropología y Poder - PiresDocumento57 pagineAntropología y Poder - PiresVanessa VillalobosNessuna valutazione finora
- Revista Istor N - 60 Historias de La PrehistoriaDocumento320 pagineRevista Istor N - 60 Historias de La PrehistoriaAna SbattellaNessuna valutazione finora
- El CeiboDocumento7 pagineEl CeiboAna SbattellaNessuna valutazione finora
- El Escuerzo Común, Un Gigante de Las PampasDocumento6 pagineEl Escuerzo Común, Un Gigante de Las PampasAna SbattellaNessuna valutazione finora
- Piensan, Sufren y Charlan Las Plantas?Documento6 paginePiensan, Sufren y Charlan Las Plantas?Ana SbattellaNessuna valutazione finora
- Mastodontes en Marcos PazDocumento4 pagineMastodontes en Marcos PazAna SbattellaNessuna valutazione finora
- Mengoni GoñalonsDocumento23 pagineMengoni GoñalonsAna SbattellaNessuna valutazione finora
- Que Es Una FalaciaDocumento6 pagineQue Es Una FalaciaAna SbattellaNessuna valutazione finora
- Huella GeneticaDocumento12 pagineHuella GeneticajhonselysNessuna valutazione finora
- Historia de La GenéticaDocumento5 pagineHistoria de La GenéticaLorena BetancourtNessuna valutazione finora
- Ácidos nucleicos: estructura y funcionesDocumento20 pagineÁcidos nucleicos: estructura y funcionesSherly Yesenia Melgarejo CastañedaNessuna valutazione finora
- Infografía de Los Tipos de Reproducción en Los Seres VivosDocumento14 pagineInfografía de Los Tipos de Reproducción en Los Seres VivosDerek ToddNessuna valutazione finora
- Guia de Biologia Noveno - 1Documento10 pagineGuia de Biologia Noveno - 1Adybeto TorresNessuna valutazione finora
- Anual SM Semana 03 - BiologíaDocumento19 pagineAnual SM Semana 03 - BiologíaGeordan SanchezNessuna valutazione finora
- Harper Bioquimica IlustradaDocumento6 pagineHarper Bioquimica Ilustradarosa zambranoNessuna valutazione finora
- Taller No. 1 Mejoramiento AnimalDocumento7 pagineTaller No. 1 Mejoramiento Animalarsened0% (2)
- PRÁCTICA #02 GENETICA Observación de Fases Del Ciclo Celular en Meristemos de Ápices Radiculares de Allium Cepa "CebollaDocumento5 paginePRÁCTICA #02 GENETICA Observación de Fases Del Ciclo Celular en Meristemos de Ápices Radiculares de Allium Cepa "CebollaRandy Jefferson ParedesNessuna valutazione finora
- 2a Micro I Clase Genética Bacteriana y THG PARTE 1 PDFDocumento43 pagine2a Micro I Clase Genética Bacteriana y THG PARTE 1 PDFSajin KomamuraNessuna valutazione finora
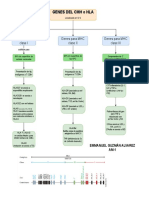
- Genes de HLA o MHCDocumento1 paginaGenes de HLA o MHCEmmanuel GNessuna valutazione finora
- Apuntes Bioquimica Udemy Maestro Camach Leanr Ingeniero en BiotecnologíaDocumento2 pagineApuntes Bioquimica Udemy Maestro Camach Leanr Ingeniero en BiotecnologíaHader CruzNessuna valutazione finora
- Importancia de La Bioquímica en La AgriculturaDocumento19 pagineImportancia de La Bioquímica en La AgriculturaJose David Aguilar Mendoza63% (8)
- Practica #05 - Mega DnaDocumento14 paginePractica #05 - Mega DnaXiory Estefany100% (1)
- PseudohermafroditaDocumento9 paginePseudohermafroditaJuliana EscuderoNessuna valutazione finora
- DR Parra-BracamonteDocumento14 pagineDR Parra-BracamonteYsik HoNessuna valutazione finora
- NGS: Secuenciación de Segunda Generación: NGS: Un Gran Concepto Codificado en Solo Tres LetrasDocumento6 pagineNGS: Secuenciación de Segunda Generación: NGS: Un Gran Concepto Codificado en Solo Tres LetrasAlvaro Martín MuñozNessuna valutazione finora
- Electroporación y Transfeccion G3Documento21 pagineElectroporación y Transfeccion G3Heidi RosalesNessuna valutazione finora
- P1 Extracción de ADN PDFDocumento6 pagineP1 Extracción de ADN PDFlauraNessuna valutazione finora
- Practica N 10 - GRUPO 1Documento12 paginePractica N 10 - GRUPO 1erikapasacheNessuna valutazione finora
- El Misterioso Celacanto Deja de Ser Un Fósil VivienteDocumento7 pagineEl Misterioso Celacanto Deja de Ser Un Fósil VivienteFERNANDO JOSE OSORIO ROSARIONessuna valutazione finora
- GametogénesisDocumento1 paginaGametogénesisgesu piero lopez mejiaNessuna valutazione finora
- Genética Bíblica 04 - Las Dos Simientes PDFDocumento7 pagineGenética Bíblica 04 - Las Dos Simientes PDFgilNessuna valutazione finora
- Genética GeneralDocumento5 pagineGenética GeneralSHIRLEYNessuna valutazione finora
- Taller de Repaso Genetica 1 - 2021Documento5 pagineTaller de Repaso Genetica 1 - 2021Yulieth Vega RosalesNessuna valutazione finora
- Clase de Repaso Modulo IVDocumento46 pagineClase de Repaso Modulo IVroostellNessuna valutazione finora
- Cuestionario de Regulación Genética.Documento19 pagineCuestionario de Regulación Genética.DanielNessuna valutazione finora
- Dogma Central de La Biologia Molecular 1-2022Documento140 pagineDogma Central de La Biologia Molecular 1-2022Andres SalgadoNessuna valutazione finora
- Escogiendo Un Vector de ClonaciónDocumento8 pagineEscogiendo Un Vector de ClonaciónFernando CidaNessuna valutazione finora
- 01 Nomenclatura de La Citogenetica HumanaDocumento18 pagine01 Nomenclatura de La Citogenetica HumanaPierina DelgadoNessuna valutazione finora