Potrebbero piacerti anche
- Istologia Ed Embriologia-Giordano PerinDocumento184 pagineIstologia Ed Embriologia-Giordano PerinGiordano Perin100% (5)
- 120 Domande FisiologiaDocumento6 pagine120 Domande FisiologiagoyshorvalNessuna valutazione finora
- Risposte Domande Esame Orale Fisiologia.Documento24 pagineRisposte Domande Esame Orale Fisiologia.Paolo Berghella100% (1)
- Enzimi PlasmaticiDocumento38 pagineEnzimi Plasmaticiapi-3704348100% (1)
- Affrontare il COVID-19 con i benefici della cannabis medicaDa EverandAffrontare il COVID-19 con i benefici della cannabis medicaNessuna valutazione finora
- AA - VV. - Compendio Di Farmacologia PDFDocumento270 pagineAA - VV. - Compendio Di Farmacologia PDFcoloromaneNessuna valutazione finora
- 4 Lezione - La Cute e I Suoi AnnessiDocumento24 pagine4 Lezione - La Cute e I Suoi AnnessiAlbert stefan PaulencoNessuna valutazione finora
- Flusso Respiratorio, O2 e CO2Documento9 pagineFlusso Respiratorio, O2 e CO2api-3704348100% (3)
- APPUNTI FisiologiaDocumento18 pagineAPPUNTI FisiologiaIrene CerutiNessuna valutazione finora
- DIARREEDocumento29 pagineDIARREEapi-3733154Nessuna valutazione finora
- (Med - Ita - Istologia Sintesi - EmbriologiaDocumento31 pagine(Med - Ita - Istologia Sintesi - EmbriologiammNessuna valutazione finora
- Muscolo Schel - Parte1Documento34 pagineMuscolo Schel - Parte1Francesco MalavoltaNessuna valutazione finora
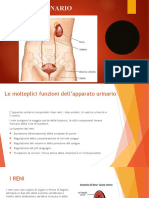
- Apparato UrinarioDocumento17 pagineApparato UrinarioCate?Nessuna valutazione finora
- Elettrocardiogramma v3Documento15 pagineElettrocardiogramma v3Marcello VecchioNessuna valutazione finora
- Metodo RuffiniDocumento11 pagineMetodo RuffiniD.DomenicoNessuna valutazione finora
- Drenaggio e Disintossicazione OmotossicologiaDocumento31 pagineDrenaggio e Disintossicazione Omotossicologiakeya Angels100% (1)
- EnzimiDocumento13 pagineEnzimiAndrea_Gerosa_3807Nessuna valutazione finora
- CALLIGARISDocumento5 pagineCALLIGARISAndreola Del Vivo100% (1)
- Gas Respiratori, Rapporto Tra Ventilazione e PerfusioneDocumento15 pagineGas Respiratori, Rapporto Tra Ventilazione e Perfusioneapi-37043480% (1)
- Equilibrio Acido Base, Fisiologia RespiratorioDocumento2 pagineEquilibrio Acido Base, Fisiologia RespiratorioVanessa SpinelliNessuna valutazione finora
- Approccio ai disordini dell'EAB: L'Equilibrio Acido-Base nella pratica clinicaDa EverandApproccio ai disordini dell'EAB: L'Equilibrio Acido-Base nella pratica clinicaNessuna valutazione finora
- Malattie Da Carenze Ed Eccessi Minerali PDFDocumento38 pagineMalattie Da Carenze Ed Eccessi Minerali PDFArmin van BurenNessuna valutazione finora
- Fisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFDocumento13 pagineFisiologia Respiratoria (Meccanica, Scambio e Trasporto Dei Gasi, Controllo Nervoso) PDFBobbyGustafson100% (1)
- Mappa Concettuale Sull'Apparato RespiratorioDocumento1 paginaMappa Concettuale Sull'Apparato RespiratorioBOLFRA100% (1)
- Apparato RespiratorioDocumento9 pagineApparato Respiratoriochiara9rossoNessuna valutazione finora
- 4b.fisiologia Apparato DigerenteDocumento9 pagine4b.fisiologia Apparato DigerenteFedericoPppzPippoNessuna valutazione finora
- Domande Compiti Fisiologia 2 - Flavio KellerDocumento33 pagineDomande Compiti Fisiologia 2 - Flavio KellerrobydiesisNessuna valutazione finora
- 1) Apparato Cardio-VascolareDocumento29 pagine1) Apparato Cardio-Vascolarecor_91Nessuna valutazione finora
- Appunti Fisiologia IDocumento27 pagineAppunti Fisiologia Ikekko88Nessuna valutazione finora
- Rimedio Per Acufeni Di Ian McCallDocumento6 pagineRimedio Per Acufeni Di Ian McCallGuadalupe HickmanNessuna valutazione finora
- ANTIOSSIDANTIDocumento13 pagineANTIOSSIDANTImodenafraNessuna valutazione finora
- 5-Stenosi MitralicaDocumento8 pagine5-Stenosi MitralicaScribdNixNessuna valutazione finora
- Respirazione CellulareDocumento29 pagineRespirazione CellulareDaniele ZigiottoNessuna valutazione finora
- 4Documento29 pagine4maria cristina ristagnoNessuna valutazione finora
- Lezioni Di Medicina Tradizionale CineseDocumento18 pagineLezioni Di Medicina Tradizionale CineseChiara BaldiniNessuna valutazione finora
- Massimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare PolmonareDocumento21 pagineMassimi Di Inspirazione, Gas Respiratori e Loro Trasporto, Velocità Di Passaggio Del Sangue Nel Capillare Polmonareapi-3704348100% (1)
- Necrosi e Autofagia CelulareDocumento86 pagineNecrosi e Autofagia Celularezolta9Nessuna valutazione finora
- 01 Muscolo ScheletricoDocumento43 pagine01 Muscolo ScheletricoaxelshippoNessuna valutazione finora
- DR Bolgan Vaccini - 09 2020 1 1Documento67 pagineDR Bolgan Vaccini - 09 2020 1 1Truth.to.be.ToldNessuna valutazione finora
- Fisiologia AppuntiDocumento20 pagineFisiologia AppuntienzocavajuoloNessuna valutazione finora
- Corso Di Anatomia TopograficaDocumento54 pagineCorso Di Anatomia TopograficaAnonymous RFVIG7IaNessuna valutazione finora
- Sistema LinfaticoDocumento9 pagineSistema Linfatico//Nessuna valutazione finora
- 4) Carboidrati - GlicolisiDocumento39 pagine4) Carboidrati - GlicolisiClaudia De LucaNessuna valutazione finora
- Apparato Respiratorio e SpirometriaDocumento9 pagineApparato Respiratorio e Spirometriaapi-3704348100% (1)
- Biochimica II - Il Ciclo Dell'Acido CitricoDocumento4 pagineBiochimica II - Il Ciclo Dell'Acido CitricoThales AlvesNessuna valutazione finora
- La Rete Dell'emergenza Sanitaria in Piemonte: Il Sistema 118Documento17 pagineLa Rete Dell'emergenza Sanitaria in Piemonte: Il Sistema 118Eumed.orgNessuna valutazione finora
- Corso Di Cannabicoltura Per PrincipiantiDocumento6 pagineCorso Di Cannabicoltura Per Principiantifoxpro123Nessuna valutazione finora
- Fisiopatologia AterosclerosiDocumento62 pagineFisiopatologia AterosclerosiLorenzo SuardiNessuna valutazione finora
- DiabeteDocumento74 pagineDiabeteapi-3863584Nessuna valutazione finora
- EMATOLOGIADocumento16 pagineEMATOLOGIAFedericoPppzPippoNessuna valutazione finora
- Vena PortaDocumento3 pagineVena Portaalessan9Nessuna valutazione finora
- Traumatologia in Terapia IntensivaDocumento25 pagineTraumatologia in Terapia IntensivaIfy NebedumNessuna valutazione finora
- Biochimica I - I LipidiDocumento4 pagineBiochimica I - I LipidiThales AlvesNessuna valutazione finora
- Apparato CircolatorioDocumento4 pagineApparato CircolatorioLuigi BorgeseNessuna valutazione finora
- Respirazione Cellulare...Documento2 pagineRespirazione Cellulare...Magda AzzilonnaNessuna valutazione finora
- Fototerapia PDFDocumento12 pagineFototerapia PDFOsteofisioNessuna valutazione finora
- Calendario SemineDocumento8 pagineCalendario SemineTua Mamma PompaNessuna valutazione finora
- Riassunti Del Tura PDFDocumento111 pagineRiassunti Del Tura PDFElena Silvia SoldeanuNessuna valutazione finora
- Linee Guida Per Pazienti Con IperomocisteinemiaDocumento32 pagineLinee Guida Per Pazienti Con Iperomocisteinemiametal76Nessuna valutazione finora
- 4a.fisiologia Apparato RespiratorioDocumento17 pagine4a.fisiologia Apparato RespiratorioFedericoPppzPippoNessuna valutazione finora
- Microbiologia IDocumento76 pagineMicrobiologia INancy LeutchNessuna valutazione finora
- 31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreDocumento6 pagine31-Anatomia II-03.03.2016-Topografia Anatomica Del CuoreSandroNessuna valutazione finora
- Immunopatologia TinaDocumento43 pagineImmunopatologia Tinabimbipulci83% (6)
- Impianto AcquaponicaDocumento27 pagineImpianto AcquaponicaSergio p. aer VALLORANI Rappresentanze impiantisticheNessuna valutazione finora
- Legumi PDFDocumento34 pagineLegumi PDFlorenzo_cavicch7255Nessuna valutazione finora
- Come concimare l’orto. Uso dei concimi organici e chimiciDa EverandCome concimare l’orto. Uso dei concimi organici e chimiciNessuna valutazione finora
- Anestetici LocaliDocumento29 pagineAnestetici LocaliMarcello VecchioNessuna valutazione finora
- ParasimpaticoliticiDocumento6 pagineParasimpaticoliticiMarcello VecchioNessuna valutazione finora
- Sistema BiliareDocumento4 pagineSistema BiliareMarcello VecchioNessuna valutazione finora
- Controllo Della CircolazioneDocumento26 pagineControllo Della CircolazioneNoemi Di FucciaNessuna valutazione finora
- Fisiopatologia FiorentinoDocumento90 pagineFisiopatologia FiorentinoMarcello VecchioNessuna valutazione finora
- ParasimpaticoliticiDocumento6 pagineParasimpaticoliticiMarcello VecchioNessuna valutazione finora
- Assimil Russo PDFDocumento295 pagineAssimil Russo PDFMarcello VecchioNessuna valutazione finora
- Guida Al PHP5Documento144 pagineGuida Al PHP5aerghNessuna valutazione finora
- Lezione 6 - Anatomia II - Organizzazione Del Grigio SpinaleDocumento8 pagineLezione 6 - Anatomia II - Organizzazione Del Grigio SpinaleMarcello VecchioNessuna valutazione finora
- Insufficienza Renale AcutaDocumento3 pagineInsufficienza Renale AcutaMarcello VecchioNessuna valutazione finora
- Lezione ArmoniaDocumento2 pagineLezione ArmoniaMarcello VecchioNessuna valutazione finora
- Guida Al PHP5Documento144 pagineGuida Al PHP5aerghNessuna valutazione finora
- Batteri MicrobiologiaDocumento4 pagineBatteri MicrobiologiaMarcello VecchioNessuna valutazione finora
- Lezione Biologia Ripasso MedicinaDocumento1 paginaLezione Biologia Ripasso MedicinaMarcello VecchioNessuna valutazione finora
- Perchè Si CredeDocumento15 paginePerchè Si CredeMarcello VecchioNessuna valutazione finora
- Batteri MicrobiologiaDocumento4 pagineBatteri MicrobiologiaMarcello VecchioNessuna valutazione finora
- BANDO Medicina 2011Documento12 pagineBANDO Medicina 2011Marcello VecchioNessuna valutazione finora
- Schmitt Eric-Emmanuel - Oscar e La Dama in RosaDocumento44 pagineSchmitt Eric-Emmanuel - Oscar e La Dama in RosaMihaela StamateNessuna valutazione finora
- Made in SediloDocumento36 pagineMade in SediloProgetto SediloNessuna valutazione finora
- 15SG2012Documento65 pagine15SG2012Linija21Nessuna valutazione finora
- Elaboró: Brenda Gutierrez ZarcoDocumento47 pagineElaboró: Brenda Gutierrez ZarcoNADIANessuna valutazione finora
- Guida Breve Di SopravvivenzaDocumento12 pagineGuida Breve Di SopravvivenzasgrispoNessuna valutazione finora
- Dizionario Italiano FriulanoDocumento18 pagineDizionario Italiano FriulanoMos CraciunNessuna valutazione finora
- Cervello, Mente e DrogheDocumento64 pagineCervello, Mente e Drogheaccasette25Nessuna valutazione finora
- Trattamento Herpes Zoster PDFDocumento28 pagineTrattamento Herpes Zoster PDFGianfranco MuntoniNessuna valutazione finora