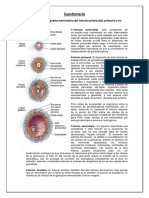
Potrebbero piacerti anche
- Aparato Reproductor FemeninoDocumento20 pagineAparato Reproductor FemeninoKoa NunchiNessuna valutazione finora
- Ovo GenesisDocumento32 pagineOvo Genesisalejandra sanchezNessuna valutazione finora
- MF0712_1 - Operaciones auxiliares en reproducción ganaderaDa EverandMF0712_1 - Operaciones auxiliares en reproducción ganaderaNessuna valutazione finora
- Ciclo Menstrual y Sus TrastornosDocumento19 pagineCiclo Menstrual y Sus TrastornosGustavo Enrique Carrasco Perez100% (1)
- Monografia Desarrollo PrenatalDocumento42 pagineMonografia Desarrollo PrenatalICASTICA33% (3)
- Ciclo Sexual ResumenDocumento3 pagineCiclo Sexual Resumenac_navarrorNessuna valutazione finora
- Guia Embriologia CeabioDocumento5 pagineGuia Embriologia CeabioHaykareth Mi Amor BelloNessuna valutazione finora
- Ciclo SexualDocumento3 pagineCiclo Sexualsponjita28Nessuna valutazione finora
- El Pibe Resumen de FecundaciónDocumento29 pagineEl Pibe Resumen de FecundaciónAntonella Violo100% (1)
- 03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesDocumento23 pagine03 Gónadas Femeninas - Origen Embriológico - Ovogénesis - Ciclos EstralesLucía María BradleyNessuna valutazione finora
- Temas de Embriologia ENURMDocumento35 pagineTemas de Embriologia ENURMLissetteAlexandraPerezNessuna valutazione finora
- Fisiologia de La FecundacionDocumento6 pagineFisiologia de La FecundacionIvan TeranNessuna valutazione finora
- Ciclo ReproductorDocumento9 pagineCiclo ReproductorMily BenincaNessuna valutazione finora
- Trabajo IvanDocumento12 pagineTrabajo IvanNatalia SáenzNessuna valutazione finora
- Resumen Embriologia LangmanDocumento13 pagineResumen Embriologia Langmanmanuel_aguirre_13Nessuna valutazione finora
- De La Ovulacion A La ImplantacionDocumento12 pagineDe La Ovulacion A La Implantacionffloresp100% (4)
- Cuestionario Cuerpo HumanoDocumento5 pagineCuestionario Cuerpo HumanoNathalia PedrozaNessuna valutazione finora
- T-6. MeiosisDocumento2 pagineT-6. MeiosisJuanJose45Nessuna valutazione finora
- ESPERMATOGÉNESISDocumento3 pagineESPERMATOGÉNESISLeo BorjasNessuna valutazione finora
- Maduracion Del Ovocito, Fecundación y Desarrollo Embrionario Equipo 2Documento65 pagineMaduracion Del Ovocito, Fecundación y Desarrollo Embrionario Equipo 2Tania GuerraNessuna valutazione finora
- Transporte de Gametos - Gineco IDocumento9 pagineTransporte de Gametos - Gineco IAndreaNessuna valutazione finora
- OvogenesisDocumento15 pagineOvogenesisRaul CuevaNessuna valutazione finora
- Ciclo MenstrualDocumento11 pagineCiclo MenstrualmelissaNessuna valutazione finora
- Primera Semana Del Desarrollo EmbrionarioDocumento33 paginePrimera Semana Del Desarrollo EmbrionarioIsabella GomezNessuna valutazione finora
- Expoccion HoyDocumento4 pagineExpoccion HoyJosé Nelson lucero huamanNessuna valutazione finora
- Embriologia Usmp SeminarioDocumento9 pagineEmbriologia Usmp SeminarioLuz Valderrama DurandNessuna valutazione finora
- GametogénesisDocumento4 pagineGametogénesisAlexander EztradaNessuna valutazione finora
- OvogénesisDocumento11 pagineOvogénesisMaria Claudia Castro HumpireNessuna valutazione finora
- Fabricio EmbrioDocumento9 pagineFabricio EmbrioFabri IbañezNessuna valutazione finora
- Ovo GenesisDocumento15 pagineOvo GenesisRosalío Mata RoblesNessuna valutazione finora
- OvogénesisDocumento3 pagineOvogénesisFernando XochitecatlNessuna valutazione finora
- Ovo GenesisDocumento14 pagineOvo GenesisJoseph K. López MolinaNessuna valutazione finora
- Fase de Crecimiento PeriantralDocumento4 pagineFase de Crecimiento PeriantralAlisNessuna valutazione finora
- Paso 2 Norida OrdoñezDocumento16 paginePaso 2 Norida OrdoñezLiliana OrdoñezNessuna valutazione finora
- Tema 1. Fisiología Del Ciclo Menstrual NormalDocumento8 pagineTema 1. Fisiología Del Ciclo Menstrual NormalcarmenNessuna valutazione finora
- Loe11 FisiologiaDocumento30 pagineLoe11 FisiologiaJuan Pablo Giraldo MontoyaNessuna valutazione finora
- Fisiología Femenina Antes Del Embarazo y Hormonas FemeninasDocumento10 pagineFisiología Femenina Antes Del Embarazo y Hormonas Femeninasnicolas100% (2)
- Primera Semana de Desarrollo EmbrionarioDocumento5 paginePrimera Semana de Desarrollo EmbrionarioEnoc GonzálezNessuna valutazione finora
- Ciclo MenstrualDocumento5 pagineCiclo MenstrualAlessandro MoralesNessuna valutazione finora
- Fisiología Femenina Antes Del Embarazo, Hormonas Femeninas y Ciclo MenstrualDocumento6 pagineFisiología Femenina Antes Del Embarazo, Hormonas Femeninas y Ciclo Menstrualjeffer665Nessuna valutazione finora
- ESPERMATOGENESISDocumento10 pagineESPERMATOGENESISAlfonso Quispe CopaNessuna valutazione finora
- OvonogenesisDocumento15 pagineOvonogenesisNicole SalamancaNessuna valutazione finora
- GAMETOGÉNESISDocumento3 pagineGAMETOGÉNESISMartina GentilNessuna valutazione finora
- Resumen Fundamentos de Estructura y FunciónDocumento43 pagineResumen Fundamentos de Estructura y FunciónGénesis JiménezNessuna valutazione finora
- Cuestionario 8Documento3 pagineCuestionario 8Victor AngelNessuna valutazione finora
- Gametogénesis y Embriología Del Aparato Genital FemeninoDocumento12 pagineGametogénesis y Embriología Del Aparato Genital FemeninoAlma Linda Peréz Santeliz100% (1)
- Meta1.3:VILLA LOPEZDocumento8 pagineMeta1.3:VILLA LOPEZFlor Denisse Villa LópezNessuna valutazione finora
- Primera Semana Del FetoDocumento14 paginePrimera Semana Del FetoJeff LNessuna valutazione finora
- CICLO OVÁRICO y EndometrialDocumento3 pagineCICLO OVÁRICO y EndometrialSelma Xiomara FragosoNessuna valutazione finora
- Reproducción Sexual 8 GADRO SEPTIEMBREDocumento6 pagineReproducción Sexual 8 GADRO SEPTIEMBREGerson OoNessuna valutazione finora
- GAMETOGÉNESISDocumento10 pagineGAMETOGÉNESISKesy PinosNessuna valutazione finora
- Espermatogenesis y OvogenesisDocumento13 pagineEspermatogenesis y OvogenesisElvis Humantupa Estrada100% (3)
- EMBRIOLOGIA GUIA ENURM Embriologia CompletoDocumento18 pagineEMBRIOLOGIA GUIA ENURM Embriologia CompletoMADDYNessuna valutazione finora
- Resumen LangmanDocumento20 pagineResumen LangmanGabrielaRojas60% (5)
- Ciclo Ovarico-FecundaciónDocumento12 pagineCiclo Ovarico-Fecundaciónkarol PonceNessuna valutazione finora
- Cuadro Comparativo Entre Ovogenesis y EspermatogenesisDocumento5 pagineCuadro Comparativo Entre Ovogenesis y Espermatogenesisgavio_ta26100% (1)
- OvogenesisDocumento15 pagineOvogenesisCatalina Rothen RodríguezNessuna valutazione finora
- Exposición Recuperación ParcialDocumento18 pagineExposición Recuperación Parcialluis AlexanderNessuna valutazione finora
- Instrumentos QuirurgicosDocumento9 pagineInstrumentos QuirurgicosDerly Serrano RuizNessuna valutazione finora
- Actividad Enzimatica Catalasa y AmilasaDocumento3 pagineActividad Enzimatica Catalasa y AmilasaDeiler HernándezNessuna valutazione finora
- Reino AnimalDocumento11 pagineReino AnimalPatricia SanchezNessuna valutazione finora
- Lectura SextoDocumento21 pagineLectura SextoDelsi Cachay VillarrealNessuna valutazione finora
- Proyecto Las BambasDocumento2 pagineProyecto Las BambasRossella Mendoza CanalesNessuna valutazione finora
- FECONIDocumento19 pagineFECONIAdriana CarranzaNessuna valutazione finora
- Capitulo 12 Rang y DaleDocumento11 pagineCapitulo 12 Rang y DaleVanessa100% (1)
- GatosDocumento9 pagineGatosGabriela Cardenas SNessuna valutazione finora
- Tema 1. Microbiologia - Dr. Gonzalez CabezaDocumento10 pagineTema 1. Microbiologia - Dr. Gonzalez CabezaChristian Rivero ValenciaNessuna valutazione finora
- Orden HymenopteraDocumento23 pagineOrden HymenopteraJHON STEVEEN CUSCUE SILVANessuna valutazione finora
- Interacciones Farmacológicas I: AntagonismoDocumento8 pagineInteracciones Farmacológicas I: AntagonismoCristobal Severo Huamani CarrionNessuna valutazione finora
- CC y PMPDocumento4 pagineCC y PMPAlejandro LluqueNessuna valutazione finora
- Machado Luis Alberto - La Revolucion de La InteligenciaDocumento58 pagineMachado Luis Alberto - La Revolucion de La InteligenciaCesar Kv Rangel100% (8)
- Team HumanaDocumento19 pagineTeam Humanamache301910Nessuna valutazione finora
- Desarrollo Del Feto Mes A MesDocumento11 pagineDesarrollo Del Feto Mes A MesMelissa LangléNessuna valutazione finora
- Pautas en La Atencion de PodologiaDocumento3 paginePautas en La Atencion de PodologiaCristofer Antonio Cañete GalazNessuna valutazione finora
- Gordimer, Nadine - Un Arma en CasaDocumento166 pagineGordimer, Nadine - Un Arma en CasaEdgar Ricardo Hernández DíazNessuna valutazione finora
- Flora y FaunaDocumento4 pagineFlora y FaunaAnonymous HbUgpL5TGNessuna valutazione finora
- Botánica, CitologíaDocumento10 pagineBotánica, CitologíaBárbara SmerkinNessuna valutazione finora
- 06 CaCu - VPH-ARDocumento1 pagina06 CaCu - VPH-ARaguirreyeni746100% (1)
- Caracterizacion Del PlatanoDocumento33 pagineCaracterizacion Del PlatanoFREDY ALEXANDER RODRIGUEZ PINEDANessuna valutazione finora
- Universidad César VallejoDocumento9 pagineUniversidad César VallejoPablo GGNessuna valutazione finora
- RECO DabrakasDocumento5 pagineRECO DabrakasLegado Pleyadiano Tecnicas HolisticasNessuna valutazione finora
- Tabla de Intercambio Alimentos, 2Documento25 pagineTabla de Intercambio Alimentos, 2Valentina AlbaNessuna valutazione finora
- Trabajo Final Evalua - Teoria - Lopez-Moya.fDocumento24 pagineTrabajo Final Evalua - Teoria - Lopez-Moya.fVANESSA LÓPEZ LÓPEZNessuna valutazione finora
- Dialnet MetodologiasDeExtraccionDeMetalesPesadosPresentesE 5628711 PDFDocumento8 pagineDialnet MetodologiasDeExtraccionDeMetalesPesadosPresentesE 5628711 PDFJorgeVictorMauriceLiraNessuna valutazione finora
- Puya de RaymundiDocumento16 paginePuya de RaymundiHedber Gamboa SantanaNessuna valutazione finora
- Guía de Observación de ActitudesDocumento10 pagineGuía de Observación de Actitudeszenovio jacomeNessuna valutazione finora
- Medio Ambient eDocumento4 pagineMedio Ambient eOverlin FernandezNessuna valutazione finora
- Informe 7 - Mendez, Forero, Pinilla-1Documento7 pagineInforme 7 - Mendez, Forero, Pinilla-1Omar Andrés BetancurNessuna valutazione finora