Potrebbero piacerti anche
- Practica PucachaDocumento1 paginaPractica PucachaJahime ArriagaNessuna valutazione finora
- RRPP PDFDocumento53 pagineRRPP PDFJhon CharlyNessuna valutazione finora
- Vega PerezDocumento108 pagineVega PerezJosé BernalNessuna valutazione finora
- Plan de Monitoreo de Metales PesadosDocumento26 paginePlan de Monitoreo de Metales PesadosJahime ArriagaNessuna valutazione finora
- Introducción y Evolución de La CalidadDocumento16 pagineIntroducción y Evolución de La CalidadJahime ArriagaNessuna valutazione finora
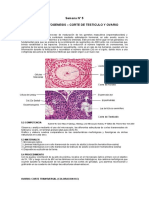
- Semana 5 PracticaDocumento2 pagineSemana 5 Practicapatricia100% (1)
- La Biología Como CienciaDocumento4 pagineLa Biología Como CienciaGustavo SantosNessuna valutazione finora
- Bioindicadores y BiomarcadoresDocumento42 pagineBioindicadores y BiomarcadoresRicardo LinaresNessuna valutazione finora
- Actividad 1 - Antecedentes HistóricosDocumento3 pagineActividad 1 - Antecedentes HistóricosAndyssyta Kessta50% (2)
- Manejo Del Estrés Hídrico en El Cultivo Del AguacateDocumento28 pagineManejo Del Estrés Hídrico en El Cultivo Del AguacateFernando Bambu de XalNessuna valutazione finora
- CarboxisomasDocumento5 pagineCarboxisomasCatalina HerreraNessuna valutazione finora
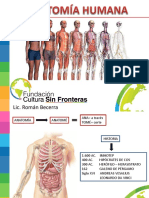
- Generalidades de Anatomia HumanaDocumento18 pagineGeneralidades de Anatomia HumanaLucas Florencio de MedeirosNessuna valutazione finora
- Tejido SanguíneoDocumento15 pagineTejido Sanguíneocarlina villamarNessuna valutazione finora
- Prez-Schultheiss 2021 Opiliones de ChileDocumento35 paginePrez-Schultheiss 2021 Opiliones de ChileCl StiebelNessuna valutazione finora
- Cambios en Estructuras Dentales y NutricionDocumento29 pagineCambios en Estructuras Dentales y NutricionNadiaDanielaDelgadoSuárezNessuna valutazione finora
- LamarckismoDocumento303 pagineLamarckismojoanNessuna valutazione finora
- PR - Respiracion Celular.Documento4 paginePR - Respiracion Celular.Margarita Moreno OrellanaNessuna valutazione finora
- LIBRO SEMILLASfi 1177140511984Documento385 pagineLIBRO SEMILLASfi 1177140511984Pedro QUNessuna valutazione finora
- Los VirusDocumento8 pagineLos VirusGenny Alejandra Jiménez MarrufoNessuna valutazione finora
- Policlínica MetropolitanaDocumento2 paginePoliclínica MetropolitanaHsiullNessuna valutazione finora
- Beneficios de La Biotecnología en Diferentes CamposDocumento10 pagineBeneficios de La Biotecnología en Diferentes CamposMiguel Torr50% (4)
- P Adaptaciones de Tallos y HojasDocumento2 pagineP Adaptaciones de Tallos y HojasABDUL HASSAN MUCHICA CHAYÑANessuna valutazione finora
- Cuerpo HumanoDocumento9 pagineCuerpo HumanoJulio Carlos Salgado BautistaNessuna valutazione finora
- TFM MEADH Cristopher Corvalan 2014Documento158 pagineTFM MEADH Cristopher Corvalan 2014Camilo PachecoNessuna valutazione finora
- Guia Sistema Reproductor Masculino 6 BasicoDocumento3 pagineGuia Sistema Reproductor Masculino 6 BasicoSUSETTENessuna valutazione finora
- Calidad de Carne de Pollo en Condiciones de Estres Ante-MortemDocumento64 pagineCalidad de Carne de Pollo en Condiciones de Estres Ante-MortemJose Garcia CruzNessuna valutazione finora
- Gametognesis BiologiaDocumento15 pagineGametognesis BiologiaJosue RebolledoNessuna valutazione finora
- Evaluación Ciencias Naturales 4 Microorganismos 7 Año 2017Documento4 pagineEvaluación Ciencias Naturales 4 Microorganismos 7 Año 2017Blanca LuisaNessuna valutazione finora
- Fisiología CardiovascularDocumento2 pagineFisiología CardiovascularImpresiones SFSNessuna valutazione finora
- 4to Año-Cromosomas Ciclo Celular Mitosis MeiosisDocumento51 pagine4to Año-Cromosomas Ciclo Celular Mitosis MeiosisSofia GomezNessuna valutazione finora
- Guia Didáctica Interacciones BiológicasDocumento11 pagineGuia Didáctica Interacciones BiológicasCecilia0% (1)
- Transfuncion Sanguínea1Documento7 pagineTransfuncion Sanguínea1Javier Sanz EstradaNessuna valutazione finora
- Modulo 2. Tejidos VegetalesDocumento45 pagineModulo 2. Tejidos VegetalesAstrid GonzalezNessuna valutazione finora
- Programa de La Materia de GenéticaDocumento14 paginePrograma de La Materia de GenéticaEnriqueRamosReaNessuna valutazione finora
- EPIGENETICA2022Documento5 pagineEPIGENETICA2022Adriana Lucero GonzalesNessuna valutazione finora