Potrebbero piacerti anche
- El GaldrabókDocumento22 pagineEl GaldrabókYo Quimica85% (41)
- Rabdovirus: RabiaDocumento29 pagineRabdovirus: RabiaMarcela Ferreira Riveros100% (1)
- Dengue FisiopatologiaDocumento65 pagineDengue FisiopatologiaNo Sbras Pero Pesadilla100% (2)
- Jabon Natural Con Aloe VeraDocumento5 pagineJabon Natural Con Aloe VeraEmanuelRic100% (1)
- Consilience La Unidad Del ConocimientoDocumento242 pagineConsilience La Unidad Del ConocimientoCarlos Isacaz100% (1)
- ADENOMATOSIS PULMONAR OVINA (Jaagsiekte)Documento12 pagineADENOMATOSIS PULMONAR OVINA (Jaagsiekte)Darwin Antezana De la RosaNessuna valutazione finora
- SilicioDocumento22 pagineSilicioJuan Diego Suárez Agualimpia100% (1)
- Cuadernillo Recortable de Lectoescritura - OrganizedDocumento25 pagineCuadernillo Recortable de Lectoescritura - OrganizedEN UN CLICKNessuna valutazione finora
- Diarrea Viral BovinaDocumento30 pagineDiarrea Viral BovinaAngie Cruz BernalNessuna valutazione finora
- RabiaDocumento29 pagineRabiaDiego Loza MNessuna valutazione finora
- Test VocacionalDocumento21 pagineTest VocacionalEfren HomNessuna valutazione finora
- Diapositivas Rabia PPT - 1890105895Documento14 pagineDiapositivas Rabia PPT - 1890105895Jaime CoronelNessuna valutazione finora
- Alvarez Alexander - Davila Peralta Ricardo - Articulo-Pátologia Aviar-10aDocumento4 pagineAlvarez Alexander - Davila Peralta Ricardo - Articulo-Pátologia Aviar-10aDayana TagleNessuna valutazione finora
- Analisis Filogenetico Del Gen de La Hemaglutinina Del Virus Distemper Canino en Perros Infectados Naturalmente en ChileDocumento27 pagineAnalisis Filogenetico Del Gen de La Hemaglutinina Del Virus Distemper Canino en Perros Infectados Naturalmente en ChileMarla ArizaNessuna valutazione finora
- Implementación de Una Técnica de Reacción en Cadena de La Polimerasa Anidada para La Detección Del Gen de La Proteína Timidin Kinasa Del Virus Herpes FelinoDocumento25 pagineImplementación de Una Técnica de Reacción en Cadena de La Polimerasa Anidada para La Detección Del Gen de La Proteína Timidin Kinasa Del Virus Herpes FelinoCARLOTA DIAZ AGUILARNessuna valutazione finora
- Universidad de Chile: Anais Sarahi Polanco VergaraDocumento9 pagineUniversidad de Chile: Anais Sarahi Polanco VergaraConstanza Pilar FigueroaNessuna valutazione finora
- SunshinevirusDocumento18 pagineSunshinevirusPilar MondragónNessuna valutazione finora
- Herpesvirus CaninoDocumento8 pagineHerpesvirus CaninoJorge Luis Martinez AmarisNessuna valutazione finora
- III Simposio Nacional de Virologia 2008 Salud UISDocumento39 pagineIII Simposio Nacional de Virologia 2008 Salud UISJeanette Prada ArismendyNessuna valutazione finora
- ArterivirusDocumento13 pagineArterivirusJocelyn LunaNessuna valutazione finora
- Apoptosis Vinculada A La Morfogénesis Del AstrovirusDocumento15 pagineApoptosis Vinculada A La Morfogénesis Del Astrovirusmiguel angelNessuna valutazione finora
- Universidad Nacional de San Martín - TarapotoDocumento35 pagineUniversidad Nacional de San Martín - Tarapotojuan jose correa ramirezNessuna valutazione finora
- MICRODocumento37 pagineMICROBRANDON YAHIR CAMACHO CARRANZANessuna valutazione finora
- Articulo Norovirus MicroDocumento10 pagineArticulo Norovirus MicroMaribel HolguinNessuna valutazione finora
- Generalidades de VirusDocumento57 pagineGeneralidades de VirusLUIS ARIZANessuna valutazione finora
- Investigacion Acerca Del Moquillo - En.esDocumento8 pagineInvestigacion Acerca Del Moquillo - En.esAlexandra GuerreroNessuna valutazione finora
- DVBDocumento4 pagineDVBJosé Hiram Sánchez GascaNessuna valutazione finora
- ResultDocumento77 pagineResultMadeline MoraNessuna valutazione finora
- Rabia: virus, estructura, patogenia e historiaDocumento35 pagineRabia: virus, estructura, patogenia e historiaXavier GarzonNessuna valutazione finora
- Diarrea ViralDocumento12 pagineDiarrea ViralSandra López JiménezNessuna valutazione finora
- Canine Distemper, Systematic ReviewDocumento39 pagineCanine Distemper, Systematic ReviewJose Luis VargasNessuna valutazione finora
- Taller Teorico - Practico de VirologiaDocumento5 pagineTaller Teorico - Practico de VirologiaANGELA NICOLE REINA BARBOSA100% (1)
- AstrovirusDocumento8 pagineAstrovirusLucía Margarita VillaNessuna valutazione finora
- Prueba Inmunocromatográfica Rápida en El Diagnóstico Del Moquillo CaninoDocumento8 paginePrueba Inmunocromatográfica Rápida en El Diagnóstico Del Moquillo CaninoemanuelagustinriveroNessuna valutazione finora
- Tecnica MolecularDocumento4 pagineTecnica MolecularCarlos reyes archundiaNessuna valutazione finora
- Trabajo de Monografia de Clinica Herpes VirusDocumento5 pagineTrabajo de Monografia de Clinica Herpes Viruskristeldelcarpio24Nessuna valutazione finora
- Evaluacion MuVDocumento6 pagineEvaluacion MuVSamir MirandaNessuna valutazione finora
- Adenovirus: receptor, replicación y clínicaDocumento3 pagineAdenovirus: receptor, replicación y clínicarodrigo RodrigoNessuna valutazione finora
- Virus Influenza: Características y AislamientoDocumento45 pagineVirus Influenza: Características y AislamientoEduardo GraciaNessuna valutazione finora
- Diagnóstico Laboratorial Del DengueDocumento45 pagineDiagnóstico Laboratorial Del DengueMaria Elena Lumbre RequeNessuna valutazione finora
- Trabajo Virologia Final 3Documento7 pagineTrabajo Virologia Final 3Claudia MedranoNessuna valutazione finora
- Tesis-Enmanuel_CespedesDocumento123 pagineTesis-Enmanuel_Cespedescristian robledoNessuna valutazione finora
- Bronquitis InfecciosaDocumento20 pagineBronquitis InfecciosaMarilys RendónNessuna valutazione finora
- Virus - MicrobiologiaDocumento28 pagineVirus - MicrobiologiaGuido Mauricio Oliva GarciaNessuna valutazione finora
- Artritis Encefalitis CaprinaDocumento2 pagineArtritis Encefalitis CaprinaAlejandria CarranzaNessuna valutazione finora
- Artículo Anaplasma Tana - En.esDocumento7 pagineArtículo Anaplasma Tana - En.esAndrés SánchezNessuna valutazione finora
- Colera PorcinoDocumento23 pagineColera PorcinoClaudia Garcia0% (1)
- Universidad Autonoma Agraria Antonio Narro: Unidad LagunaDocumento27 pagineUniversidad Autonoma Agraria Antonio Narro: Unidad LagunaZulliseuidy Samantha Franco MuñizNessuna valutazione finora
- Informe 9 de ParasitoDocumento15 pagineInforme 9 de ParasitoGarito95Nessuna valutazione finora
- Infecciones TropicalesDocumento33 pagineInfecciones Tropicaleswilber ramos condoriNessuna valutazione finora
- Peritonitis Infecciosa FelinaDocumento27 paginePeritonitis Infecciosa FelinaJair SaavedraNessuna valutazione finora
- virologia porcinaDocumento11 paginevirologia porcinaGustavo SteinbregerNessuna valutazione finora
- Gammaherpesvirus y Hepadnavirus FelinosDocumento4 pagineGammaherpesvirus y Hepadnavirus Felinosherediaruben1896Nessuna valutazione finora
- Tarea 12 VirologíaDocumento22 pagineTarea 12 VirologíaBertha Felix TarazonaNessuna valutazione finora
- 3 Síndrome Reproductivo y Respiratorio Porcino IIDocumento6 pagine3 Síndrome Reproductivo y Respiratorio Porcino IIAxoncomunicacionNessuna valutazione finora
- Rotavirus Virol.Documento33 pagineRotavirus Virol.Lucho Zafra HaroNessuna valutazione finora
- Dialnet AspectosMolecularesDelVirusDeLaParvovirosisCaninaY 4943854 PDFDocumento9 pagineDialnet AspectosMolecularesDelVirusDeLaParvovirosisCaninaY 4943854 PDFAna CauichNessuna valutazione finora
- Aglutinación en Látex, Elisa y HemoaglutinaciónDocumento7 pagineAglutinación en Látex, Elisa y HemoaglutinaciónKate GarcíaNessuna valutazione finora
- Resumen BovinosDocumento2 pagineResumen BovinosPao Gisell LeonNessuna valutazione finora
- Parotiditis y RotavirusDocumento38 pagineParotiditis y RotavirusLidia Samaniego CazorlaNessuna valutazione finora
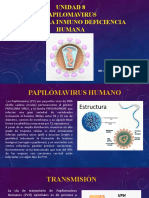
- Unidad 8 Papilomavirus - VihDocumento18 pagineUnidad 8 Papilomavirus - VihLISBETH PANIAGUA GUTIERREZNessuna valutazione finora
- Parvovirinae - ParvovirusDocumento16 pagineParvovirinae - ParvovirusJhoel soto machaccaNessuna valutazione finora
- Virus de La Fiebre AftosaDocumento4 pagineVirus de La Fiebre AftosaRocio CardenasNessuna valutazione finora
- 1.-Taxones A Estudiar: Tema 16: Virus Arnmc (+) Envueltos-Desnudos Y RetrovirusDocumento9 pagine1.-Taxones A Estudiar: Tema 16: Virus Arnmc (+) Envueltos-Desnudos Y RetrovirusJuan Rodriguez PérezNessuna valutazione finora
- Tratamiento de Coronavirus Felino Con El Análogo de NucleósidoDocumento23 pagineTratamiento de Coronavirus Felino Con El Análogo de NucleósidoCarolinaNessuna valutazione finora
- Ancylostoma CaninumDocumento1 paginaAncylostoma CaninumRosita Vera TallesNessuna valutazione finora
- DX DiabetesDocumento9 pagineDX DiabetesRosita Vera TallesNessuna valutazione finora
- PeriodontitisDocumento5 paginePeriodontitisRosita Vera TallesNessuna valutazione finora
- Justificación para El Uso de Jaulas para Las Aves de CrianzaDocumento4 pagineJustificación para El Uso de Jaulas para Las Aves de CrianzaRosita Vera TallesNessuna valutazione finora
- Historia Clinica Orientada A ProblemasDocumento14 pagineHistoria Clinica Orientada A ProblemasFranz HuamaniNessuna valutazione finora
- DistemperDocumento8 pagineDistemperlokitotristeNessuna valutazione finora
- Manual de prácticas de la asignatura Práctica de medicina de perrosDocumento34 pagineManual de prácticas de la asignatura Práctica de medicina de perrosStephanie Toxiic ToOxNessuna valutazione finora
- Manual de prácticas de la asignatura Práctica de medicina de perrosDocumento34 pagineManual de prácticas de la asignatura Práctica de medicina de perrosStephanie Toxiic ToOxNessuna valutazione finora
- DistemperDocumento8 pagineDistemperlokitotristeNessuna valutazione finora
- 30 Medicina GatosDocumento38 pagine30 Medicina GatosConstanza Contreras CastroNessuna valutazione finora
- LeishmaniaDocumento12 pagineLeishmaniaRosita Vera TallesNessuna valutazione finora
- AntisépticosDocumento5 pagineAntisépticosRosita Vera TallesNessuna valutazione finora
- Interpretacion Radiografica PDFDocumento10 pagineInterpretacion Radiografica PDFEdgar Ricardo Anastacio NavarroNessuna valutazione finora
- ToracotomíaDocumento49 pagineToracotomíaRosita Vera TallesNessuna valutazione finora
- Radiologia PDFDocumento45 pagineRadiologia PDFRosita Vera TallesNessuna valutazione finora
- Aislamiento de Esplenocitos de RatónDocumento2 pagineAislamiento de Esplenocitos de RatónRosita Vera TallesNessuna valutazione finora
- IntroducciónDocumento12 pagineIntroducciónRosita Vera TallesNessuna valutazione finora
- RadiodiagnósticoDocumento7 pagineRadiodiagnósticoRosita Vera TallesNessuna valutazione finora
- Enterolobium CyclocarpumDocumento4 pagineEnterolobium CyclocarpumRosita Vera TallesNessuna valutazione finora
- Anatomía del aparato reproductor macho ovinoDocumento6 pagineAnatomía del aparato reproductor macho ovinoRosita Vera Talles50% (2)
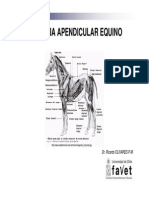
- Miologia Apendicular Equino USTDocumento30 pagineMiologia Apendicular Equino USTzuzuki220291Nessuna valutazione finora
- Repartido Digestion en Reticulo RumenDocumento30 pagineRepartido Digestion en Reticulo RumenMauro D FNessuna valutazione finora
- Birna Viri DaeDocumento10 pagineBirna Viri DaeRosita Vera TallesNessuna valutazione finora
- RotavirusDocumento8 pagineRotavirusRosita Vera TallesNessuna valutazione finora
- Antisepticos Mecanismo AcciónDocumento15 pagineAntisepticos Mecanismo AcciónAndrea Urra FuenzalidaNessuna valutazione finora
- Anatomia Del MachoDocumento14 pagineAnatomia Del MachoRosita Vera TallesNessuna valutazione finora
- Atlas A Color de Patologia Porcina PDFDocumento191 pagineAtlas A Color de Patologia Porcina PDFtiburonpirata100% (7)
- Concept o Medio Ambient eDocumento29 pagineConcept o Medio Ambient eRosita Vera TallesNessuna valutazione finora
- Cap 1Documento33 pagineCap 1Rosita Vera TallesNessuna valutazione finora
- Evidencia 1Documento6 pagineEvidencia 1juancarloscostales0% (2)
- Quiz-Escenario 3: 75/75 en 11 minutosDocumento8 pagineQuiz-Escenario 3: 75/75 en 11 minutosMariat Borrero RomeroNessuna valutazione finora
- Estructuras celularesDocumento2 pagineEstructuras celularesCruz cibelys LucianoNessuna valutazione finora
- Factores ambientales y su influencia en el trabajoDocumento20 pagineFactores ambientales y su influencia en el trabajoLuis Fernando SanchezNessuna valutazione finora
- TFR Miranda Gran CanariaDocumento10 pagineTFR Miranda Gran CanariaLeticia DorbessanNessuna valutazione finora
- Beneficios de Los JabonesDocumento5 pagineBeneficios de Los Jabonesglory cifreNessuna valutazione finora
- Piel HidratadaDocumento1 paginaPiel HidratadaLizzett Gutiérrez GarcíaNessuna valutazione finora
- CRONOGRAMA PARA ASISTENTES y PUBLICO EN GENERALDocumento8 pagineCRONOGRAMA PARA ASISTENTES y PUBLICO EN GENERALYampier LopezNessuna valutazione finora
- Guia 5 C. Naturales Quinto PDFDocumento3 pagineGuia 5 C. Naturales Quinto PDFsebas204Nessuna valutazione finora
- Practica 3 MicrosDocumento6 paginePractica 3 MicrosARIADNA NATASHA MANYA MORANNessuna valutazione finora
- 62 ASIGNATURA Manejo de Fauna Silvestre SEPTDocumento10 pagine62 ASIGNATURA Manejo de Fauna Silvestre SEPTNahara Hdz MontejoNessuna valutazione finora
- Tema 3 NeuriDocumento46 pagineTema 3 Neuriyungmario MRNessuna valutazione finora
- Guia de Alergia para Residentes y Atencion PrimariaDocumento300 pagineGuia de Alergia para Residentes y Atencion PrimariaMarco Dominguez100% (1)
- Taller de Replicacion y TraduccionDocumento4 pagineTaller de Replicacion y TraduccionNataliaCharryNessuna valutazione finora
- 25 Lecturas para Mejorar y Controlar La Velocidad Lectora Parte1Documento10 pagine25 Lecturas para Mejorar y Controlar La Velocidad Lectora Parte1MaríaJoséVásquezdeLópezNessuna valutazione finora
- Guía Lectura Critica 7 S.PDocumento10 pagineGuía Lectura Critica 7 S.Pclaudia yolima lopezNessuna valutazione finora
- Caja Entomología Clasificación de InsectosDocumento22 pagineCaja Entomología Clasificación de InsectosDenisNessuna valutazione finora
- MC 0067857Documento423 pagineMC 0067857german hernandezNessuna valutazione finora
- Ejercicios de Concentracin de Energa Vita 1Documento2 pagineEjercicios de Concentracin de Energa Vita 1FERNANDO OUBIÑANessuna valutazione finora
- MercadeoDocumento34 pagineMercadeoBrayant Stick SANCHEZ BOLANOSNessuna valutazione finora
- UROANALISIS SedimentoDocumento14 pagineUROANALISIS SedimentoRosa SantosNessuna valutazione finora
- Resumen Vias BiliaresDocumento7 pagineResumen Vias BiliaresMOCHABATYNessuna valutazione finora
- Práctica Análisis Heces 2022Documento9 paginePráctica Análisis Heces 2022ERIKABIOANALISISNessuna valutazione finora
- U1 - Actividad 1. La Base Química de La VidaDocumento4 pagineU1 - Actividad 1. La Base Química de La VidaLupe PescadorNessuna valutazione finora