Potrebbero piacerti anche
- Respuestas Khor CognicionDocumento17 pagineRespuestas Khor CognicionMariel Valadez70% (10)
- 1º ESo Biología Tema 1-Anaya A.C.Documento13 pagine1º ESo Biología Tema 1-Anaya A.C.MinervaNessuna valutazione finora
- DESCARGAR LEER DOWNLOAD READ. Descripción. Anatomía de Los Animales Domésticos. Tomo I PDF - Descargar, Leer ENGLISH VERSIONDocumento10 pagineDESCARGAR LEER DOWNLOAD READ. Descripción. Anatomía de Los Animales Domésticos. Tomo I PDF - Descargar, Leer ENGLISH VERSIONPedro Germain Gutierrez Vergara100% (1)
- Ensayo Sobre El Sistema Nervioso de Los AnimalesDocumento3 pagineEnsayo Sobre El Sistema Nervioso de Los Animalesvioleta silva75% (4)
- Práctica de Equilibrio de Hardy-WeinbergDocumento3 paginePráctica de Equilibrio de Hardy-WeinbergLUIS DANIEL G�MEZ PEREIRA100% (1)
- Mindful LivingDocumento15 pagineMindful LivingLucero ProfetaNessuna valutazione finora
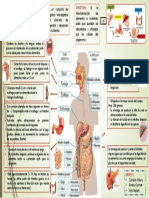
- Infografia Del Sistema DigestivoDocumento1 paginaInfografia Del Sistema DigestivoLuis E De Abreu57% (7)
- Obbi AperetiDocumento36 pagineObbi AperetiLuis Omo Elegua100% (4)
- Adobe Scan 10 Jul 2023Documento5 pagineAdobe Scan 10 Jul 2023Luzmila Lucia Apaza YanaricoNessuna valutazione finora
- 1.0.reconocemos Nuestro Desarrollo Histórico de La Biología.4abc - Fv. Lic. Karen Reyes PDFDocumento6 pagine1.0.reconocemos Nuestro Desarrollo Histórico de La Biología.4abc - Fv. Lic. Karen Reyes PDFREBECA REYES QUISPENessuna valutazione finora
- Icfes 11 1p 2018Documento3 pagineIcfes 11 1p 2018Carlos Hernandez RodriguezNessuna valutazione finora
- Homeostasis A Nivel Planetario.Documento8 pagineHomeostasis A Nivel Planetario.KAREN ROJAS75% (4)
- Ordenamiento Ecologico Territorial de La Costa de SinaloaDocumento214 pagineOrdenamiento Ecologico Territorial de La Costa de SinaloaLisl EcheverriNessuna valutazione finora
- Protesis Parcial Fija 2 Delmy ReyesDocumento3 pagineProtesis Parcial Fija 2 Delmy ReyesDelmy ReyesNessuna valutazione finora
- Aceites para El CabelloDocumento4 pagineAceites para El Cabellojean callesNessuna valutazione finora
- Informe #7 - Bioca Sem - G04Documento9 pagineInforme #7 - Bioca Sem - G04Yordi CuzqueNessuna valutazione finora
- Práctica 3. Verificación y Cuantificación AdnDocumento4 paginePráctica 3. Verificación y Cuantificación AdnCristina FajardoNessuna valutazione finora
- Cuadernillo de Actividades Extracción y Purificación de Ácidos NucleicosDocumento4 pagineCuadernillo de Actividades Extracción y Purificación de Ácidos NucleicosJeremy David Nenjer BahamondeNessuna valutazione finora
- Taller Biología Noveno AdnDocumento4 pagineTaller Biología Noveno AdnSamuel GonzalezNessuna valutazione finora
- Revee, Capitulo 2Documento9 pagineRevee, Capitulo 2Abril CollerNessuna valutazione finora
- T1. Retículo Endoplásmico LisoDocumento9 pagineT1. Retículo Endoplásmico LisoAokima SanNessuna valutazione finora
- ECTOPARASITOSDocumento5 pagineECTOPARASITOSBárbara MansillaNessuna valutazione finora
- PROTEÍNASDocumento15 paginePROTEÍNASDavid ArzuzaNessuna valutazione finora
- GRUPO 43 Tarea 4 - Metabolismo Grupo - 43Documento26 pagineGRUPO 43 Tarea 4 - Metabolismo Grupo - 43yeris selena mora marroquin33% (3)
- Neuropsicologia de La LecturaDocumento29 pagineNeuropsicologia de La LecturaMargoth NaviaNessuna valutazione finora
- Trabajo Unidad 2 HigieneDocumento32 pagineTrabajo Unidad 2 Higienesandra castañedaNessuna valutazione finora
- Aislamiento de Rhizobium SPDocumento3 pagineAislamiento de Rhizobium SPFabioAGM11Nessuna valutazione finora
- Taller No. 1Documento4 pagineTaller No. 1saray castilloNessuna valutazione finora
- Estrategias de Reciclaje Arquitectónico PDFDocumento163 pagineEstrategias de Reciclaje Arquitectónico PDFgescobarg4377Nessuna valutazione finora
- Parque Nacional SangayDocumento3 pagineParque Nacional SangayCristhian Alvear100% (1)