Potrebbero piacerti anche
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Wanda Wajang Purwa R Soetrisno 2 27Documento13 pagineWanda Wajang Purwa R Soetrisno 2 27BobbyGunarsoNessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Barata Yudha Sunardi DMDocumento72 pagineBarata Yudha Sunardi DMBobbyGunarsoNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5795)
- À Å° Ó Ðåä - Sonnet of FountainDocumento5 pagineÀ Å° Ó Ðåä - Sonnet of FountainBobbyGunarsoNessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Kesehatan Dan Keadilan Sosial inDocumento20 pagineKesehatan Dan Keadilan Sosial inBobbyGunarsoNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Metrics and Evaluation: Metric Sample Calculation Advantages DisadvantagesDocumento10 pagineMetrics and Evaluation: Metric Sample Calculation Advantages DisadvantagesBobbyGunarsoNessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (345)
- Sebuah Patung Singa Dari Candi Prambanan Diapit Dengan Dua Kapataru Dan KinaraDocumento28 pagineSebuah Patung Singa Dari Candi Prambanan Diapit Dengan Dua Kapataru Dan KinaraBobbyGunarsoNessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Development of Cell-Cycle Inhibitors For Cancer Therapy: M.A. Dickson and G.K. SchwartzDocumento8 pagineDevelopment of Cell-Cycle Inhibitors For Cancer Therapy: M.A. Dickson and G.K. SchwartzBobbyGunarsoNessuna valutazione finora
- 9781603841948Documento1 pagina9781603841948BobbyGunarsoNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Kirim Ukai HipertensiDocumento18 pagineKirim Ukai HipertensiBobbyGunarsoNessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Modul Matrikulasi FTSF - Bu BetiDocumento146 pagineModul Matrikulasi FTSF - Bu BetiBobbyGunarsoNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Water Density: Published On Healing EarthDocumento2 pagineWater Density: Published On Healing EarthBobbyGunarsoNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Indonesia Raya For PianoDocumento5 pagineIndonesia Raya For PianoBobbyGunarso50% (2)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Modul UKAI 2015 FarmakokimiaDocumento41 pagineModul UKAI 2015 FarmakokimiaBobbyGunarsoNessuna valutazione finora
- Three Kingdoms RevisitedDocumento152 pagineThree Kingdoms RevisitedBobbyGunarsoNessuna valutazione finora
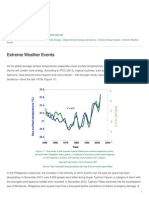
- Healing Earth - Extreme Weather Events - 2015-04-21Documento2 pagineHealing Earth - Extreme Weather Events - 2015-04-21BobbyGunarsoNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Indonesia Raya For Symphony OrchestraDocumento13 pagineIndonesia Raya For Symphony OrchestraBobbyGunarsoNessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- I Believe in Springtime - Rutter - EnglishDocumento3 pagineI Believe in Springtime - Rutter - EnglishBobbyGunarsoNessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Growth and DevelopmentDocumento47 pagineGrowth and DevelopmentMark Anthony FloresNessuna valutazione finora
- 2010 PSPM Kedah Biology 123 W AnsDocumento93 pagine2010 PSPM Kedah Biology 123 W Ansjee2kk100% (1)
- Physiology Su00Documento42 paginePhysiology Su00Mahad Ahmed HilowleNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- Basic Science Note Primary FourDocumento16 pagineBasic Science Note Primary Foursulaiman mohammedNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1091)
- Noise and Vibration HazardsDocumento28 pagineNoise and Vibration HazardsAKANKSHANessuna valutazione finora
- Butterflies and Their Contribution in Ecosystem: A ReviewDocumento4 pagineButterflies and Their Contribution in Ecosystem: A ReviewMuhammadBarrylAnggriawanNessuna valutazione finora
- Economics 11Documento2 pagineEconomics 11KevinB CaigasNessuna valutazione finora
- BMB Technical Bulletin No 2016-10 Outline of Cave Wetland Management PlanDocumento4 pagineBMB Technical Bulletin No 2016-10 Outline of Cave Wetland Management PlanAnonymous dtceNuyIFI100% (1)
- Diabetes Mellitus 1Documento96 pagineDiabetes Mellitus 1manideepreddyNessuna valutazione finora
- Dna WorksheetDocumento8 pagineDna WorksheetLuis100% (6)
- 1 s2.0 S2667376222000130 MainDocumento8 pagine1 s2.0 S2667376222000130 MainelfiahNessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Jurnal KelDocumento3 pagineJurnal Kelandre erlisNessuna valutazione finora
- Clinical NeurodynamicsDocumento41 pagineClinical Neurodynamicsramesh2007-mptNessuna valutazione finora
- Tumor Monocyte Content Predicts ImmunochemotherapyDocumento28 pagineTumor Monocyte Content Predicts ImmunochemotherapyDi V.Nessuna valutazione finora
- Water Treatment With Exceptional Virus Inactivation Using Activated Carbon Modified With Silver Ag and Copper Oxide CuO Nanoparticles PDFDocumento13 pagineWater Treatment With Exceptional Virus Inactivation Using Activated Carbon Modified With Silver Ag and Copper Oxide CuO Nanoparticles PDFMarcela FernandesNessuna valutazione finora
- Barrett's EsophagusDocumento10 pagineBarrett's EsophagusaryadroettninguNessuna valutazione finora
- Bio Lab 2 RespirationDocumento7 pagineBio Lab 2 RespirationDominic100% (1)
- Week 1 - Hematology 2 LectureDocumento8 pagineWeek 1 - Hematology 2 LectureRubenne Miles ElagasNessuna valutazione finora
- Bio 101 LAB EXER 1Documento24 pagineBio 101 LAB EXER 1Alliza Andal100% (1)
- Active Site of EnzymesDocumento11 pagineActive Site of EnzymesVictor AkinseyeNessuna valutazione finora
- Adolescence 8th Edition: Chapter One: Biological TransitionsDocumento35 pagineAdolescence 8th Edition: Chapter One: Biological TransitionsNick BuzaNessuna valutazione finora
- The Body Container A New Perspective On The Body EgoDocumento21 pagineThe Body Container A New Perspective On The Body EgoClo Mn100% (1)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Unit 6 Growth and Development and Care of The AdolescentDocumento17 pagineUnit 6 Growth and Development and Care of The AdolescentKyla CapituloNessuna valutazione finora
- Photosynthesis - WikipediaDocumento22 paginePhotosynthesis - Wikipediaمحمد جانNessuna valutazione finora
- Pro-Bio Beads: Solgar Vitamin and HerbDocumento2 paginePro-Bio Beads: Solgar Vitamin and HerbM Usman KhanNessuna valutazione finora
- UG Prospectus 2014 - Full VersionDocumento197 pagineUG Prospectus 2014 - Full VersionLina Syazana Md RadziNessuna valutazione finora
- Melamine in FoodsDocumento15 pagineMelamine in FoodsAnura Senarathna100% (4)
- A Compilation of Philippine Environmental LawsDocumento39 pagineA Compilation of Philippine Environmental LawsRussel Sirot100% (3)
- Septal Extension Graft ArticlesDocumento7 pagineSeptal Extension Graft Articlesjose orlando sturla pelleranoNessuna valutazione finora
- Child Development An Introduction Santrock 14th Edition Test BankDocumento60 pagineChild Development An Introduction Santrock 14th Edition Test BankWilliam Williams100% (36)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDa EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionValutazione: 4 su 5 stelle4/5 (404)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDa EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityValutazione: 4 su 5 stelle4/5 (34)