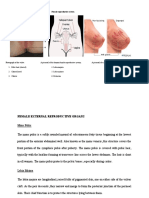
Potrebbero piacerti anche
- The Second G SpotDocumento27 pagineThe Second G SpotJhonatan HinestrozaNessuna valutazione finora
- BarCharts QuickStudy Reproductive SystemDocumento2 pagineBarCharts QuickStudy Reproductive SystemRUDY792100% (4)
- How to Give Her Mind-Blowing Oral PleasureDocumento64 pagineHow to Give Her Mind-Blowing Oral Pleasuresolo subo mierd100% (1)
- Anatomy Physiology of The Male Female Reproductive SystemDocumento17 pagineAnatomy Physiology of The Male Female Reproductive SystemLilian JerotichNessuna valutazione finora
- Female Reproductive System PDFDocumento48 pagineFemale Reproductive System PDFTeppy the GreatNessuna valutazione finora
- Anatomy and PhysiologyDocumento8 pagineAnatomy and PhysiologyG3-PORTILLA John MartNessuna valutazione finora
- Scrotum Anatomy and Scanning TechniqueDocumento7 pagineScrotum Anatomy and Scanning TechniqueAnonymous wyF4yQZwZNessuna valutazione finora
- Anatomy Physiology of Female Reproductive SystemDocumento96 pagineAnatomy Physiology of Female Reproductive SystemRabin BasnetNessuna valutazione finora
- Human Reproduction PDFDocumento35 pagineHuman Reproduction PDFsqhaa100% (4)
- Reproductive SystemDocumento6 pagineReproductive Systemsubi100% (1)
- Notes On The Human Female Reproductive SystemDocumento10 pagineNotes On The Human Female Reproductive Systemfichte121Nessuna valutazione finora
- Investigatory Project of BiologyDocumento18 pagineInvestigatory Project of BiologyRanjana Singh87% (31)
- Female Reproductive System Anatomy and PhysiologyDocumento5 pagineFemale Reproductive System Anatomy and PhysiologyIkea Balhon100% (3)
- Anatomy of Female Reproductive SystemDocumento7 pagineAnatomy of Female Reproductive SystemseongeokNessuna valutazione finora
- Ovarian Cyst FinalDocumento26 pagineOvarian Cyst FinalReicy Aquino SacularNessuna valutazione finora
- WEEK 3 Day 1 Lesson 11: Define Menstrual CycleDocumento10 pagineWEEK 3 Day 1 Lesson 11: Define Menstrual Cyclejoice dela cruz100% (1)
- Maternal and Child Health NursingDocumento6 pagineMaternal and Child Health NursingRoanne DivinagraciaNessuna valutazione finora
- Understanding Hysterectomy: A Guide to the Surgical ProcedureDocumento22 pagineUnderstanding Hysterectomy: A Guide to the Surgical ProcedureGinger Enireht HTaibNessuna valutazione finora
- Anatomy and Physiology of Female Reproductive SystemDocumento6 pagineAnatomy and Physiology of Female Reproductive SystemFarlyn Real MoralesNessuna valutazione finora
- Obstetrics, Gynaecology-Textbook of Obstetrics and Gynaecology For Medical Students, 2nd Edition-Akin Agboola-2006Documento559 pagineObstetrics, Gynaecology-Textbook of Obstetrics and Gynaecology For Medical Students, 2nd Edition-Akin Agboola-2006fagiy51113Nessuna valutazione finora
- Female Reproductive System OverviewDocumento49 pagineFemale Reproductive System OverviewJohaifha CompaniaNessuna valutazione finora
- Anatomy and PhysiologyDocumento8 pagineAnatomy and PhysiologyAironne ManguilimotanNessuna valutazione finora
- Anatomy and PhysiologyDocumento14 pagineAnatomy and PhysiologyIvee BustaNessuna valutazione finora
- Reproductive Anatomy and Physiology: BY DR: AhmedDocumento82 pagineReproductive Anatomy and Physiology: BY DR: AhmedAhmed ElryahNessuna valutazione finora
- Female RSystemDocumento71 pagineFemale RSystemicliftonguytonNessuna valutazione finora
- REWIEW OF FEMALE REPRODUCTIVE SYSTEM NewDocumento49 pagineREWIEW OF FEMALE REPRODUCTIVE SYSTEM NewnamitaNessuna valutazione finora
- 12b. Reproduction Female ReproductionDocumento16 pagine12b. Reproduction Female ReproductionNishant ShahNessuna valutazione finora
- Study of the Female Reproductive SystemDocumento6 pagineStudy of the Female Reproductive SystemShreeya SharmaNessuna valutazione finora
- Anatomy and PhysiologyDocumento5 pagineAnatomy and Physiologychinchin ramosNessuna valutazione finora
- Female Reproductive SystemDocumento7 pagineFemale Reproductive SystemjonaNessuna valutazione finora
- Nursing Process for C-Section RecoveryDocumento8 pagineNursing Process for C-Section Recoverymarkie917Nessuna valutazione finora
- Anatomy and Physiology of Female Reproductive System:: VaginaDocumento5 pagineAnatomy and Physiology of Female Reproductive System:: VaginaphallicNessuna valutazione finora
- Written by AHMED ATEF K-719610990 AHMED KHALIL K-719610923: Anatomy Assignment 2Documento15 pagineWritten by AHMED ATEF K-719610990 AHMED KHALIL K-719610923: Anatomy Assignment 2Dr. Mohammad JamaliNessuna valutazione finora
- Female RSsystem 2Documento90 pagineFemale RSsystem 2icliftonguytonNessuna valutazione finora
- 2021.2 - Exam Q&ADocumento203 pagine2021.2 - Exam Q&ALucas Victor AlmeidaNessuna valutazione finora
- Uterus AnatomyDocumento7 pagineUterus AnatomyJoseph RadislaoNessuna valutazione finora
- Introduction to Women's Health: An Overview of the Female Reproductive System and Menstrual CycleDocumento47 pagineIntroduction to Women's Health: An Overview of the Female Reproductive System and Menstrual CycleDarren LimNessuna valutazione finora
- Congenital Abnormalities of The UterusDocumento7 pagineCongenital Abnormalities of The Uterusمحمود الموسويNessuna valutazione finora
- Reproductive System HandoutDocumento21 pagineReproductive System HandoutJei SanNessuna valutazione finora
- Anatomy and Physiology: Female External GenitaliaDocumento85 pagineAnatomy and Physiology: Female External GenitaliaJonah Rei BeatoNessuna valutazione finora
- Ovaries & Female Reproductive OrgansDocumento2 pagineOvaries & Female Reproductive OrgansFrancez Anne GuanzonNessuna valutazione finora
- Human Body Flat Design Presentation Pink VariantDocumento16 pagineHuman Body Flat Design Presentation Pink VariantAngie MejaritoNessuna valutazione finora
- Anatomy and Physiology - EctopicDocumento4 pagineAnatomy and Physiology - EctopicReyzelinNessuna valutazione finora
- Female Reproductive System (Human)Documento21 pagineFemale Reproductive System (Human)RiazAhmedNessuna valutazione finora
- Anatomy of the Cervix and its PhysiologyDocumento8 pagineAnatomy of the Cervix and its PhysiologyNicaMariannePañaNessuna valutazione finora
- Lecture Note On Female Reproductive System by Miss FranciscaDocumento10 pagineLecture Note On Female Reproductive System by Miss FranciscachinecheremoledimmaNessuna valutazione finora
- Anatomy and PhysiologyDocumento6 pagineAnatomy and Physiologybadgurl08Nessuna valutazione finora
- Screenshot 2022-03-28 at 10.17.47 PMDocumento127 pagineScreenshot 2022-03-28 at 10.17.47 PMqdvvqvxn7gNessuna valutazione finora
- Anatomy and Physiology of Female Reproductive System: External Generative OrgansDocumento28 pagineAnatomy and Physiology of Female Reproductive System: External Generative OrgansAlmira Gamao100% (1)
- Female Reproductive SystemDocumento7 pagineFemale Reproductive SystemyannieNessuna valutazione finora
- Female Reproductive SystemDocumento6 pagineFemale Reproductive SystemAlyanna L. ArquillanoNessuna valutazione finora
- Anatomy and Physiology Reproductive System: ComponentsDocumento7 pagineAnatomy and Physiology Reproductive System: ComponentsMark Nel Nuñez100% (1)
- Human ReproductionDocumento96 pagineHuman Reproductionsanya razaNessuna valutazione finora
- Female Reproductive AnatomyDocumento7 pagineFemale Reproductive AnatomyRalph_John_Pac_8337Nessuna valutazione finora
- Femalereproductivesystem 120329125448 Phpapp02Documento18 pagineFemalereproductivesystem 120329125448 Phpapp02Crapster GamingNessuna valutazione finora
- Anatomy and PhysiologyDocumento7 pagineAnatomy and PhysiologyKristine Alejandro100% (1)
- Female Reproductive System HistologyDocumento9 pagineFemale Reproductive System HistologyzaidNessuna valutazione finora
- Cattle Reproducive SystemDocumento12 pagineCattle Reproducive SystemSurajNessuna valutazione finora
- Female Reproductive SystemDocumento11 pagineFemale Reproductive SystemArianna Jasmine MabungaNessuna valutazione finora
- Reproductive Organ in MammalsDocumento11 pagineReproductive Organ in MammalstajudeenabdullahaNessuna valutazione finora
- Female Reproductive SystemDocumento17 pagineFemale Reproductive SystemShefali GoyalNessuna valutazione finora
- Anatomy of the Female Reproductive OrgansDocumento2 pagineAnatomy of the Female Reproductive OrgansJahn Dela RosaNessuna valutazione finora
- Enumerate The Parts and Discuss Its Function For The Following Organs/system of Farm AnimalsDocumento27 pagineEnumerate The Parts and Discuss Its Function For The Following Organs/system of Farm AnimalsJaryll Vhasti P LagumbayNessuna valutazione finora
- Female Reproductive System Anatomy and PhysiologyDocumento5 pagineFemale Reproductive System Anatomy and PhysiologyGeevee Naganag VentulaNessuna valutazione finora
- UterusDocumento3 pagineUterusBernadette FlorendoNessuna valutazione finora
- Cow ReproductiveDocumento5 pagineCow Reproductiveastyrafe100% (1)
- Anatomy and Physiology CompleteDocumento9 pagineAnatomy and Physiology CompleteJohn BretanaNessuna valutazione finora
- Lesson 8 Week 10 The Sexual Self Reproductive System by Sir CSDDocumento10 pagineLesson 8 Week 10 The Sexual Self Reproductive System by Sir CSDEdgardo IlaoNessuna valutazione finora
- The female reproductive system explainedDocumento29 pagineThe female reproductive system explainedRuben James A. PecsonNessuna valutazione finora
- Sperm Check Fertility Testing GuideDocumento4 pagineSperm Check Fertility Testing GuideMyHomeTestingNessuna valutazione finora
- Ovary JumpingDocumento4 pagineOvary JumpingTess SixNessuna valutazione finora
- A Standardized Classification of Hypospadias: Marek OrkiszewskiDocumento5 pagineA Standardized Classification of Hypospadias: Marek OrkiszewskibenimhepfiNessuna valutazione finora
- Cow ReproductiveDocumento5 pagineCow Reproductiveastyrafe100% (1)
- L6 - Anatomy of Female Reproductive SystemDocumento60 pagineL6 - Anatomy of Female Reproductive SystemJIA YUET NGNessuna valutazione finora
- Penis: Abdomen Skin CircumcisionDocumento4 paginePenis: Abdomen Skin CircumcisionTatadarz Auxtero LagriaNessuna valutazione finora
- The Parts and The Functions of The Female Reproductive SystemDocumento40 pagineThe Parts and The Functions of The Female Reproductive SystemLeo John OctavianoNessuna valutazione finora
- Juanpablo Miguel ZAVALETA RODRIGUEZ - Human ReproductionDocumento23 pagineJuanpablo Miguel ZAVALETA RODRIGUEZ - Human ReproductionJuanpablo Miguel ZAVALETA RODRIGUEZNessuna valutazione finora
- BDS MCQSDocumento4 pagineBDS MCQSMuhammad kamran ameerNessuna valutazione finora
- Female and Male Reproductive System DiagramsDocumento4 pagineFemale and Male Reproductive System DiagramsHanna AsmadNessuna valutazione finora
- An Evolution of Aesthetic Gynaecology-1Documento4 pagineAn Evolution of Aesthetic Gynaecology-1SANTOSH HOSPITALNessuna valutazione finora
- Human Reproduction - The Complete ProcessDocumento13 pagineHuman Reproduction - The Complete ProcessMahendhiran MariappanNessuna valutazione finora
- Scie10 - Binded - SY 2021-2022 - Weeks 1 To 4Documento38 pagineScie10 - Binded - SY 2021-2022 - Weeks 1 To 4Shin EscuadroNessuna valutazione finora
- Teaching LessonDocumento13 pagineTeaching LessonRizky AmaliaNessuna valutazione finora
- Chapter 16 -- Human Sexual Reproductive SystemDocumento8 pagineChapter 16 -- Human Sexual Reproductive SystemChaw Wei HengNessuna valutazione finora
- 2.reproduction in Lower and Higher Animals 1Documento29 pagine2.reproduction in Lower and Higher Animals 1nileshmpharmNessuna valutazione finora
- Antepartal Care: Prepared By: Shalini Santiago-Barroso, RN MANDocumento11 pagineAntepartal Care: Prepared By: Shalini Santiago-Barroso, RN MANJabelle Mae DoteNessuna valutazione finora
- Reproduction in Human BeingsDocumento6 pagineReproduction in Human BeingsanjupalNessuna valutazione finora