Potrebbero piacerti anche
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- CSEC Integrated Science June 1995 P1Documento7 pagineCSEC Integrated Science June 1995 P1Saintm101Nessuna valutazione finora
- Medicinal PlantsDocumento151 pagineMedicinal Plantsallanquartermaine0% (1)
- Medicinal PlantsDocumento151 pagineMedicinal Plantsallanquartermaine0% (1)
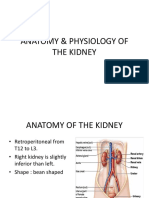
- Anatomy & Physiology of KidneyDocumento13 pagineAnatomy & Physiology of KidneyEmira EzunaNessuna valutazione finora
- (2002 Wentzinger DKK) Inhibition of Squalene Synthase and Squalene Epoxidase in Tobacco Cells Triggers An Up-Regulation of 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase PDFDocumento13 pagine(2002 Wentzinger DKK) Inhibition of Squalene Synthase and Squalene Epoxidase in Tobacco Cells Triggers An Up-Regulation of 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase PDFArinta Purwi SuhartiNessuna valutazione finora
- Aaaaaa KKDocumento1 paginaAaaaaa KKArinta Purwi SuhartiNessuna valutazione finora
- 497 FullDocumento5 pagine497 FullArinta Purwi SuhartiNessuna valutazione finora
- Molecules 15 00040Documento18 pagineMolecules 15 00040Arinta Purwi SuhartiNessuna valutazione finora
- (2012 Ye & Bhatia) REVIEW - Metabolic Engineering For The Production of Clinically Important Molecules - Omega-3 Fatty Acids, Artemisinin, and TaxolDocumento14 pagine(2012 Ye & Bhatia) REVIEW - Metabolic Engineering For The Production of Clinically Important Molecules - Omega-3 Fatty Acids, Artemisinin, and TaxolArinta Purwi SuhartiNessuna valutazione finora
- Antimicrobial Activity of Crude Extracts 2Documento6 pagineAntimicrobial Activity of Crude Extracts 2Arinta Purwi SuhartiNessuna valutazione finora
- NPH 12325Documento17 pagineNPH 12325Arinta Purwi SuhartiNessuna valutazione finora
- (2009, Anthony DKK) Optimization of The Mevalonate-Based Isoprenoid Biosynthetic in E Coli For Production Amorpha-4,11-DieneDocumento7 pagine(2009, Anthony DKK) Optimization of The Mevalonate-Based Isoprenoid Biosynthetic in E Coli For Production Amorpha-4,11-DieneArinta Purwi SuhartiNessuna valutazione finora
- (2004, Bailey DKK) Prediction of Anti-Plasmodial Activity of A Annua - Application of 1H NMR Spectroscopy and ChemometricsDocumento10 pagine(2004, Bailey DKK) Prediction of Anti-Plasmodial Activity of A Annua - Application of 1H NMR Spectroscopy and ChemometricsArinta Purwi SuhartiNessuna valutazione finora
- (2013 Pu DKK) Expression and Localization of Amorpha-4,11-Diene Synthase in Artemisia Annua L.Documento6 pagine(2013 Pu DKK) Expression and Localization of Amorpha-4,11-Diene Synthase in Artemisia Annua L.Arinta Purwi SuhartiNessuna valutazione finora
- KhawajaDocumento4 pagineKhawajaArinta Purwi SuhartiNessuna valutazione finora
- 03 ADocumento10 pagine03 AArinta Purwi SuhartiNessuna valutazione finora
- IJAMSCR 14 205 SreenagavalliDocumento3 pagineIJAMSCR 14 205 SreenagavalliArinta Purwi SuhartiNessuna valutazione finora
- FS Physio Babies and Toddlers Mar 13Documento7 pagineFS Physio Babies and Toddlers Mar 13Arinta Purwi SuhartiNessuna valutazione finora
- Antibiotic Treatment For Cystic FibrosisDocumento102 pagineAntibiotic Treatment For Cystic Fibrosischar1356Nessuna valutazione finora
- Journal Pone 0068231Documento8 pagineJournal Pone 0068231bhatubimNessuna valutazione finora
- Finding Out About Cystic Fibrosis - A Guide For Parents: Factsheet - March 2013Documento22 pagineFinding Out About Cystic Fibrosis - A Guide For Parents: Factsheet - March 2013Arinta Purwi SuhartiNessuna valutazione finora
- Ni Hms 135180Documento19 pagineNi Hms 135180Arinta Purwi SuhartiNessuna valutazione finora
- MeleDocumento4 pagineMeleArinta Purwi SuhartiNessuna valutazione finora
- HarrDocumento11 pagineHarrArinta Purwi SuhartiNessuna valutazione finora
- Ni Hms 135180Documento19 pagineNi Hms 135180Arinta Purwi SuhartiNessuna valutazione finora
- Antimicrobial Activity of Crude Extracts 2Documento6 pagineAntimicrobial Activity of Crude Extracts 2Arinta Purwi SuhartiNessuna valutazione finora
- The Pathogenesis and Pathophysiology of Type 1 and PDFDocumento12 pagineThe Pathogenesis and Pathophysiology of Type 1 and PDFasmawatiNessuna valutazione finora
- 5 Ajps 3 1 2013Documento4 pagine5 Ajps 3 1 2013Arinta Purwi SuhartiNessuna valutazione finora
- Making CosmeticsDocumento9 pagineMaking CosmeticsBogdanNessuna valutazione finora
- Spell Bee 2023-24Documento3 pagineSpell Bee 2023-24Ayush DubeyNessuna valutazione finora
- BPM 1 Spring 2019 FTM Module - Lecture and DLA Objectives Term 1 Professionalism ObjectivesDocumento11 pagineBPM 1 Spring 2019 FTM Module - Lecture and DLA Objectives Term 1 Professionalism ObjectivesNoelani-Mei Ascio50% (2)
- 34 AsfiksiaDocumento40 pagine34 AsfiksiaDaeng Anggit AdirahmanNessuna valutazione finora
- IB DP Biology 1.3 Membrane Structure QuestionDocumento15 pagineIB DP Biology 1.3 Membrane Structure QuestionikaNessuna valutazione finora
- Male and Female Genital Systems 2Documento30 pagineMale and Female Genital Systems 2negmm2226Nessuna valutazione finora
- Normal Test ReportDocumento2 pagineNormal Test ReportBilal RashidNessuna valutazione finora
- Rapidchem National Dealers Plist April 2023Documento2 pagineRapidchem National Dealers Plist April 2023lifesaviourdyenNessuna valutazione finora
- Class 10 Biology Chemistry and Physics Extra QuestionsDocumento4 pagineClass 10 Biology Chemistry and Physics Extra QuestionsSankalp KharayatNessuna valutazione finora
- OMM Block 1 Lab Study GuideDocumento18 pagineOMM Block 1 Lab Study Guidejoe doweNessuna valutazione finora
- Which Elements Are Essential For Human LifeDocumento9 pagineWhich Elements Are Essential For Human LifeJordon AlvaradoNessuna valutazione finora
- Grade 11 Biology The EyeDocumento5 pagineGrade 11 Biology The EyeDexter TorringtonNessuna valutazione finora
- Mtle - Hema 1Documento50 pagineMtle - Hema 1Leogene Earl FranciaNessuna valutazione finora
- Biology Human Reproduction PDFDocumento15 pagineBiology Human Reproduction PDFmbsureshNessuna valutazione finora
- Respiratory SystemDocumento3 pagineRespiratory SystemUzumaki Naruto (NARUTADIKS)Nessuna valutazione finora
- AnatomyDocumento23 pagineAnatomyMiracle For NursesNessuna valutazione finora
- Physioex Exercise 2 Activity 5Documento5 paginePhysioex Exercise 2 Activity 5Hanifa NabilaNessuna valutazione finora
- (Ha Lab) Sas#9Documento9 pagine(Ha Lab) Sas#9Erwin RomeroNessuna valutazione finora
- Antaomy of GITDocumento5 pagineAntaomy of GITMike GNessuna valutazione finora
- Anti AgingDocumento6 pagineAnti AgingZubair Akram JanjuaNessuna valutazione finora
- Qdoc - Tips Biology Project On Immunity For Class 12thDocumento25 pagineQdoc - Tips Biology Project On Immunity For Class 12thPadma VsNessuna valutazione finora
- Radio Study of Middle EarDocumento44 pagineRadio Study of Middle EarDOC AJNessuna valutazione finora
- Low Level Light Therapy For The Aging Face - July 2016 Clinics in Plastic SurgeryDocumento10 pagineLow Level Light Therapy For The Aging Face - July 2016 Clinics in Plastic SurgeryarexixNessuna valutazione finora
- The Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchDocumento13 pagineThe Thermoregulatory Theory of Yawning: What We Know From Over 5 Years of ResearchAndré AmorimNessuna valutazione finora
- Foundation - Module Guide 2023,1st Yr MbbsDocumento31 pagineFoundation - Module Guide 2023,1st Yr Mbbshudafatimasyeda77Nessuna valutazione finora
- Mcqs On Excretory SystemDocumento3 pagineMcqs On Excretory SystemIshfaq LoneNessuna valutazione finora
- Botany Plant Anatomy SEM II H SEM I GDocumento34 pagineBotany Plant Anatomy SEM II H SEM I GMartha-Angella MassaquoiNessuna valutazione finora
- Psyn 102 ReviewerDocumento3 paginePsyn 102 ReviewerP Harvey PuducayNessuna valutazione finora
- IGCSE - Bio - Lesson Plan 6 - CoordinationDocumento3 pagineIGCSE - Bio - Lesson Plan 6 - CoordinationHisokagen100% (2)