Potrebbero piacerti anche
- Corneal ReflexDocumento12 pagineCorneal Reflexgerald_azarconNessuna valutazione finora
- Binoc SummationDocumento5 pagineBinoc SummationwillemlrouxNessuna valutazione finora
- Pupillary Light ReflexDocumento2 paginePupillary Light ReflexJohn OsborneNessuna valutazione finora
- Physiology of Suppression in Strabismic Amblyopia: PerspectiveDocumento5 paginePhysiology of Suppression in Strabismic Amblyopia: Perspectiveimran_india12Nessuna valutazione finora
- Interocular suppression in amblyopia delays and attenuates hemodynamic responseDocumento12 pagineInterocular suppression in amblyopia delays and attenuates hemodynamic responseYuke PutriNessuna valutazione finora
- ReflexesDocumento6 pagineReflexeskristie_25100% (1)
- The Pupillary Light Reflex: How Our Eyes Adjust to LightDocumento7 pagineThe Pupillary Light Reflex: How Our Eyes Adjust to LightGian RafananNessuna valutazione finora
- Visual-Vestibular Interaction - Basic Science To Clinical RelevanceDocumento13 pagineVisual-Vestibular Interaction - Basic Science To Clinical RelevanceEnrico ArmatoNessuna valutazione finora
- Preprint. Szpak et al., Beyond Feeling Sick -The Visual and Cognitive Aftereffects of Virtual RealityDocumento9 paginePreprint. Szpak et al., Beyond Feeling Sick -The Visual and Cognitive Aftereffects of Virtual Realitytimothy.dchrist18Nessuna valutazione finora
- Eye Journal PonkopDocumento7 pagineEye Journal PonkopIndra PermanaNessuna valutazione finora
- BINOCULAR VISION LAB EXAMINES STEREOSCOPY AND RIVALRYDocumento11 pagineBINOCULAR VISION LAB EXAMINES STEREOSCOPY AND RIVALRYDr-Chufo IzquierdoNessuna valutazione finora
- Stimulus Xicker Alters Interocular Grouping During Binocular RivalryDocumento7 pagineStimulus Xicker Alters Interocular Grouping During Binocular RivalrysethNessuna valutazione finora
- Is All The Asthenopia SameDocumento8 pagineIs All The Asthenopia SameYaltiNessuna valutazione finora
- ProP Neurologic+ExaminationDocumento6 pagineProP Neurologic+ExaminationAna Paula Sabino VoltoliniNessuna valutazione finora
- Pupil Reflex Test ExplainedDocumento11 paginePupil Reflex Test ExplainedHenry Jonathan BadongenNessuna valutazione finora
- Abnormality of Eyeball PositionDocumento23 pagineAbnormality of Eyeball PositionAlexNessuna valutazione finora
- Convergence: Group 2Documento29 pagineConvergence: Group 2Kwenzie FortalezaNessuna valutazione finora
- How Brain Makes Worlds Tab Lei 0387Documento4 pagineHow Brain Makes Worlds Tab Lei 0387dbach108Nessuna valutazione finora
- Mod1 BinoDocumento2 pagineMod1 BinoNady BalberoNessuna valutazione finora
- Sulykos Brain Res 2011Documento8 pagineSulykos Brain Res 2011sofia_nakouNessuna valutazione finora
- NervoussystemlabreportDocumento9 pagineNervoussystemlabreportapi-296594736Nessuna valutazione finora
- Abnormal Fixational Eye Movements in Amblyopia: PurposeDocumento17 pagineAbnormal Fixational Eye Movements in Amblyopia: PurposeLinda MutiaraNessuna valutazione finora
- Seeing The World Dimly: The Impact of Early Visual Deficits On Visual Experience in SchizophreniaDocumento10 pagineSeeing The World Dimly: The Impact of Early Visual Deficits On Visual Experience in SchizophreniaLilianneNessuna valutazione finora
- Binocular Rivalry: An Introductory ExplorationDocumento19 pagineBinocular Rivalry: An Introductory ExplorationyyuioyyuiuuNessuna valutazione finora
- PupilgameDocumento55 paginePupilgameBataa BattulgaNessuna valutazione finora
- Neurology & Pupils (No 467)Documento231 pagineNeurology & Pupils (No 467)Mohamed GaberNessuna valutazione finora
- Physio Lab PBLDocumento4 paginePhysio Lab PBLtheodore_estradaNessuna valutazione finora
- Saccadic Behavior during the Response to Pure Disparity Vergence StimuliDocumento11 pagineSaccadic Behavior during the Response to Pure Disparity Vergence StimuliMadhumithaMahadevanNessuna valutazione finora
- CH 7 - Visual Acuity, Geometric Optical Effects of Spectacles, and Aniseikonia, P. 114-123-EmailDocumento10 pagineCH 7 - Visual Acuity, Geometric Optical Effects of Spectacles, and Aniseikonia, P. 114-123-EmailAmi MisiciNessuna valutazione finora
- Asymmetry of BlinkingDocumento15 pagineAsymmetry of BlinkingHugo JunkersNessuna valutazione finora
- Pupillary Light Reflex - Wikipedia - Https - en - Wikipedia.org - Wiki - Pupillary - Light - ReflexDocumento6 paginePupillary Light Reflex - Wikipedia - Https - en - Wikipedia.org - Wiki - Pupillary - Light - ReflexMatthew PrestonNessuna valutazione finora
- Unconscious Processing of Direct Gaze Revealed by ERP StudyDocumento8 pagineUnconscious Processing of Direct Gaze Revealed by ERP StudyJohnNessuna valutazione finora
- Lab 03 Reflexes Referred PainDocumento14 pagineLab 03 Reflexes Referred PainViraj TatikondaNessuna valutazione finora
- MYDRIATIC AND MIOTIC EFFECTSDocumento6 pagineMYDRIATIC AND MIOTIC EFFECTSManik BashaNessuna valutazione finora
- Binocular VisionDocumento49 pagineBinocular VisionekaNessuna valutazione finora
- Pupil: Photopupillary ReflexDocumento24 paginePupil: Photopupillary ReflexAyra Nakrish Alvarez100% (3)
- Latent Stereopsis For Motion in Depth in Strabismic AmblyopiaDocumento11 pagineLatent Stereopsis For Motion in Depth in Strabismic AmblyopiaYuke PutriNessuna valutazione finora
- What Methods Did We Employ in This Experiment?: Blind SpotDocumento5 pagineWhat Methods Did We Employ in This Experiment?: Blind SpotvlopezNessuna valutazione finora
- OCULOPATHY - Disproves the orthodox and theoretical bases upon which glasses are so freely prescribed, and puts forward natural remedial methods of treatment for what are sometimes termed incurable visual defectsDa EverandOCULOPATHY - Disproves the orthodox and theoretical bases upon which glasses are so freely prescribed, and puts forward natural remedial methods of treatment for what are sometimes termed incurable visual defectsNessuna valutazione finora
- Pupillary Reflex and AccomodationDocumento2 paginePupillary Reflex and AccomodationVincent Lau Bi ShengNessuna valutazione finora
- Chapter 8 Ocular Motor ControlDocumento1 paginaChapter 8 Ocular Motor ControlDeena Al NuaimiNessuna valutazione finora
- Chapter 2 - Heterophoria, Pages 15-40Documento26 pagineChapter 2 - Heterophoria, Pages 15-40blueice100% (1)
- Clinical Exam for MS SignsDocumento2 pagineClinical Exam for MS SignsssNessuna valutazione finora
- Seeing Sound: How Auditory Stimuli Can Alter Visual PerceptionDocumento21 pagineSeeing Sound: How Auditory Stimuli Can Alter Visual PerceptionSofronis ZafeiriadisNessuna valutazione finora
- Worth's Three Degrees of FusionDocumento1 paginaWorth's Three Degrees of FusionPham Son0% (1)
- Visual Acuity Toddler TestDocumento21 pagineVisual Acuity Toddler TestDimsyafelNessuna valutazione finora
- Pupil Respose AssessmentDocumento4 paginePupil Respose AssessmentRrichard Prieto MmallariNessuna valutazione finora
- Accommodative and Nonaccommodative Treatment of PresbyopiaDocumento2 pagineAccommodative and Nonaccommodative Treatment of PresbyopiaZega AgustianNessuna valutazione finora
- Ocular dominance columns visual stimulationDocumento6 pagineOcular dominance columns visual stimulationAlex MNessuna valutazione finora
- tmp275B TMPDocumento7 paginetmp275B TMPFrontiersNessuna valutazione finora
- اخطاء انكسار 2Documento3 pagineاخطاء انكسار 2Hassan AljaberiNessuna valutazione finora
- Chapter 10 Intentional Motor Disorders and The ApraxiasDocumento20 pagineChapter 10 Intentional Motor Disorders and The ApraxiasPPDSNeuroUnsri RSMHNessuna valutazione finora
- Eye Dxtics and Eye DisordersDocumento53 pagineEye Dxtics and Eye DisordersJanine Dela CruzNessuna valutazione finora
- 16-Amedi Et Al Neuron05Documento14 pagine16-Amedi Et Al Neuron05Daniel FloresNessuna valutazione finora
- 10 - Accommodation & Pupillary Light ReflexDocumento46 pagine10 - Accommodation & Pupillary Light ReflexSugumar YathavanNessuna valutazione finora
- Eye Pupillary Dilation and ConstrictionDocumento13 pagineEye Pupillary Dilation and Constrictionmusaib0831Nessuna valutazione finora
- Parameters Right Eye (Control) Left Eye (Test) : Effect of Drug On Rabbit'S Eye by CalDocumento4 pagineParameters Right Eye (Control) Left Eye (Test) : Effect of Drug On Rabbit'S Eye by CalSai tejendraNessuna valutazione finora
- 106 - CH 22 - Anomalies of Convergence and Divergence, P. 500-507Documento8 pagine106 - CH 22 - Anomalies of Convergence and Divergence, P. 500-507Francisco Vicent PachecoNessuna valutazione finora
- Neurological ExaminationDocumento23 pagineNeurological ExaminationMuhammad Fahmy100% (1)
- The Structure of A Kidney NephronDocumento4 pagineThe Structure of A Kidney NephronMarie St. Louis100% (1)
- Vision and MemoryDocumento10 pagineVision and MemoryMarie St. LouisNessuna valutazione finora
- EpitheliumDocumento10 pagineEpitheliumMarie St. Louis100% (1)
- Org Paper Draft-1scribdDocumento2 pagineOrg Paper Draft-1scribdMarie St. LouisNessuna valutazione finora
- Vision Lecture IIDocumento12 pagineVision Lecture IIMarie St. LouisNessuna valutazione finora
- Eye Memory VisonDocumento15 pagineEye Memory VisonMarie St. LouisNessuna valutazione finora
- US Internal Revenue Service: f2106 AccessibleDocumento2 pagineUS Internal Revenue Service: f2106 AccessibleIRS100% (1)
- Vision Powerpoint IDocumento22 pagineVision Powerpoint IMarie St. Louis100% (1)
- Org Paper Draft-1scribdDocumento2 pagineOrg Paper Draft-1scribdMarie St. LouisNessuna valutazione finora
- msl302 L11a Motivating Soldiers SlidesDocumento16 paginemsl302 L11a Motivating Soldiers SlidesMarie St. LouisNessuna valutazione finora
- Vision LectureDocumento24 pagineVision LectureMarie St. Louis100% (1)
- HowDocumento8 pagineHowMarie St. LouisNessuna valutazione finora
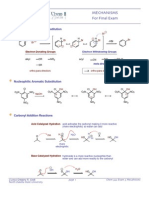
- Mechanisms For Final Exam: Electrophilic Aromatic SubstitutionDocumento4 pagineMechanisms For Final Exam: Electrophilic Aromatic SubstitutionMarie St. LouisNessuna valutazione finora
- msl302 l11c Consideration of Others SlidesDocumento44 paginemsl302 l11c Consideration of Others SlidesMarie St. LouisNessuna valutazione finora
- Summary of Organic ReactionsDocumento21 pagineSummary of Organic ReactionsMarie St. LouisNessuna valutazione finora
- Organic Chemistry Final Exam KeyDocumento10 pagineOrganic Chemistry Final Exam KeybrookNessuna valutazione finora
- EyeDocumento3 pagineEyeMarie St. LouisNessuna valutazione finora
- Finalreview 03Documento30 pagineFinalreview 03Marie St. LouisNessuna valutazione finora
- NMR 13c 05Documento34 pagineNMR 13c 05Marie St. LouisNessuna valutazione finora
- Organic Chem Reactions Mindmap PDFDocumento7 pagineOrganic Chem Reactions Mindmap PDFgsbabil100% (1)
- Organic Chemistry 342 ReactionsDocumento11 pagineOrganic Chemistry 342 ReactionsFrancisco QuirozNessuna valutazione finora
- Letter To Creditor For Reduction of DebtDocumento4 pagineLetter To Creditor For Reduction of DebtMarie St. Louis0% (1)
- Cell DivisionDocumento8 pagineCell DivisionMarie St. Louis100% (1)
- Debora Biomass and Alternative FuelDocumento58 pagineDebora Biomass and Alternative FuelMarie St. LouisNessuna valutazione finora
- Sexual HarrassementDocumento10 pagineSexual HarrassementMarie St. LouisNessuna valutazione finora
- Water PowerpointDocumento16 pagineWater PowerpointMarie St. LouisNessuna valutazione finora
- Restriction Enzyme LabDocumento12 pagineRestriction Enzyme LabMarie St. LouisNessuna valutazione finora
- Cells of Erogenous Zones Powerpoint FinalDocumento32 pagineCells of Erogenous Zones Powerpoint FinalMarie St. Louis100% (1)
- Res Enzyme Lab IIDocumento2 pagineRes Enzyme Lab IIMarie St. LouisNessuna valutazione finora
- ENDEC Letter Head BlankDocumento5 pagineENDEC Letter Head BlankklynchelleNessuna valutazione finora
- Full Download Cost Management A Strategic Emphasis 8th Edition Blocher Solutions ManualDocumento35 pagineFull Download Cost Management A Strategic Emphasis 8th Edition Blocher Solutions Manualrassyacidaq100% (37)
- Design Example Report: Title Specification Application Author Document Number Date RevisionDocumento22 pagineDesign Example Report: Title Specification Application Author Document Number Date RevisionEibar016Nessuna valutazione finora
- Broadband FTTX Ordering Guide CO-108716Documento12 pagineBroadband FTTX Ordering Guide CO-108716Gustavo MoriceNessuna valutazione finora
- Space 22Documento148 pagineSpace 22danae olivaresNessuna valutazione finora
- Capitol Steel Corporation-RSB PDFDocumento1 paginaCapitol Steel Corporation-RSB PDFeg choloNessuna valutazione finora
- DAGR BrochureDocumento2 pagineDAGR BrochureNasir Khan BhittaniNessuna valutazione finora
- Tech Manual Index PDFDocumento36 pagineTech Manual Index PDFMartín Ignacio del CampoNessuna valutazione finora
- PR 2 Module 2Documento18 paginePR 2 Module 2Earl Laurence PlatinoNessuna valutazione finora
- PSACA Presents: The Philly Cheesesteak Round 1 TossupsDocumento11 paginePSACA Presents: The Philly Cheesesteak Round 1 TossupsGhnl NglgrnlrNessuna valutazione finora
- Method 8310 Polynuclear Aromatic Hydrocarbons (PAH) HPLCDocumento13 pagineMethod 8310 Polynuclear Aromatic Hydrocarbons (PAH) HPLCridermateNessuna valutazione finora
- Slideshare Sample Essays SPM 2013Documento30 pagineSlideshare Sample Essays SPM 2013Adheera D-raNessuna valutazione finora
- Test Unit 2Documento3 pagineTest Unit 2RAMONA SECUNessuna valutazione finora
- MacLean Kevin J 200712 MScEngDocumento200 pagineMacLean Kevin J 200712 MScEngGiuseppe TizzaniNessuna valutazione finora
- Effective Warranty for Caterpillar EnginesDocumento2 pagineEffective Warranty for Caterpillar EnginesMartin McTaggartNessuna valutazione finora
- Introduction To Systems Thinking Tools - EngDocumento55 pagineIntroduction To Systems Thinking Tools - EngDan JecanNessuna valutazione finora
- Fastener TheoryDocumento4 pagineFastener TheoryAnonymous OFP2ygPIdNessuna valutazione finora
- AXIS Camera Station S1148 Recorder: User ManualDocumento39 pagineAXIS Camera Station S1148 Recorder: User ManualfguerreroaNessuna valutazione finora
- Assisted Reproductive Technology (ART)Documento6 pagineAssisted Reproductive Technology (ART)Rabelais MedinaNessuna valutazione finora
- Dramatic Prefiguration in Plato's RepublicDocumento10 pagineDramatic Prefiguration in Plato's RepublicJames JoyceNessuna valutazione finora
- Guideline HEGDocumento5 pagineGuideline HEGMushlihani HaniNessuna valutazione finora
- Lab 3-Section a-6410710052 ทัชพรDocumento12 pagineLab 3-Section a-6410710052 ทัชพรทัชรพร ปรีชาNessuna valutazione finora
- Download Novela canaima de romulo gallegos descargarDocumento3 pagineDownload Novela canaima de romulo gallegos descargarMariel sarahyNessuna valutazione finora
- ComputerDocumento16 pagineComputerdeepshooters BabuNessuna valutazione finora
- Dwnload Full Chemical Engineering Design 2nd Edition Towler Solutions Manual PDFDocumento35 pagineDwnload Full Chemical Engineering Design 2nd Edition Towler Solutions Manual PDFandrewduffydckjxfspnq100% (10)
- Baterías AlphaDocumento2 pagineBaterías Alphadisanty94Nessuna valutazione finora
- Base Isolation SystemDocumento8 pagineBase Isolation SystemIsiaka AbibuNessuna valutazione finora
- Elements of Remote SensingDocumento9 pagineElements of Remote Sensingsaamy100% (3)
- Daftar Perusahaan Top Indonesia Beserta Alamat Dan EmailnyaDocumento174 pagineDaftar Perusahaan Top Indonesia Beserta Alamat Dan EmailnyaRomeo Must Die100% (1)
- UntitledDocumento1 paginaUntitledPedro MiguelNessuna valutazione finora