Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
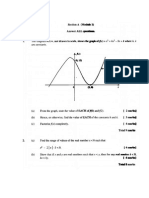
- PBL2Documento12 paginePBL2brown_chocolate87643Nessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- 17th Jan 2014 II SEM Abdomen Lecture ObjectivesDocumento3 pagine17th Jan 2014 II SEM Abdomen Lecture Objectivesbrown_chocolate87643Nessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- 14th Jan 2014 Gross Anatomy Abdomen Lecture Objectives-3Documento2 pagine14th Jan 2014 Gross Anatomy Abdomen Lecture Objectives-3brown_chocolate87643Nessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Cardiac CycleDocumento38 pagineCardiac CycleDeepika KapoorNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (400)
- Principles of PharmacologyDocumento11 paginePrinciples of Pharmacologybrown_chocolate87643Nessuna valutazione finora
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Basic Human Pathology Lecture #9 Vascular HemodynamicsDocumento45 pagineBasic Human Pathology Lecture #9 Vascular Hemodynamicsqadi78Nessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Severity of Acute Stress Symptoms Child Age 11 To 17Documento3 pagineSeverity of Acute Stress Symptoms Child Age 11 To 17brown_chocolate87643Nessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- 2014 Report Estimates of Diabetes and Its Burden in The United StatesDocumento8 pagine2014 Report Estimates of Diabetes and Its Burden in The United Statesbrown_chocolate87643Nessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- DSM V - TocDocumento9 pagineDSM V - Tocjv10gmailNessuna valutazione finora
- Meningitis, Encephalitis and Brain Abscess - Aaims - HBD - IV-1Documento72 pagineMeningitis, Encephalitis and Brain Abscess - Aaims - HBD - IV-1brown_chocolate87643Nessuna valutazione finora
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- Cape MCQ 08-13Documento51 pagineCape MCQ 08-13brown_chocolate87643Nessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (74)
- Cape Yearly Unit 1 98-05Documento61 pagineCape Yearly Unit 1 98-05brown_chocolate87643Nessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Cape Yearly Unit 1 98-05Documento61 pagineCape Yearly Unit 1 98-05brown_chocolate87643Nessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Mathcapeunit 1&2 2011-2006qDocumento92 pagineMathcapeunit 1&2 2011-2006qbrown_chocolate87643Nessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Normocytic Anemia'sDocumento14 pagineNormocytic Anemia'sbrown_chocolate87643Nessuna valutazione finora
- 2 Test For GlucoseDocumento4 pagine2 Test For GlucoseAcer MasterNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- POM Class 1 For MD7-2Documento2 paginePOM Class 1 For MD7-2brown_chocolate87643Nessuna valutazione finora
- Question 3 Micro B-2Documento3 pagineQuestion 3 Micro B-2brown_chocolate87643Nessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Phil Must Be Asked For Permission To Sample Blood For A HCV and HIV Test. If He Refuses Then It Must Be Assumed He Is A Carrier of The VirusDocumento3 paginePhil Must Be Asked For Permission To Sample Blood For A HCV and HIV Test. If He Refuses Then It Must Be Assumed He Is A Carrier of The Virusbrown_chocolate87643Nessuna valutazione finora
- Summer Course 2014 FlyerDocumento1 paginaSummer Course 2014 Flyerbrown_chocolate87643Nessuna valutazione finora
- Nimesulide Patients Still Exposed To A Risk of Severe HepatitisDocumento2 pagineNimesulide Patients Still Exposed To A Risk of Severe Hepatitisbrown_chocolate87643Nessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Real Comm Studies IA-2Documento7 pagineReal Comm Studies IA-2brown_chocolate87643Nessuna valutazione finora
- Focus For Final Exams Spring 2015Documento1 paginaFocus For Final Exams Spring 2015brown_chocolate87643Nessuna valutazione finora
- AAIMS INVITATION - 4th Anniversary LectureDocumento1 paginaAAIMS INVITATION - 4th Anniversary Lecturebrown_chocolate87643Nessuna valutazione finora
- Introduction JoviintroDocumento4 pagineIntroduction Joviintrobrown_chocolate87643Nessuna valutazione finora
- Ghouls Bhowling Alley Kiss of Death Wiked Cricket Grab Bag and More!!!Documento2 pagineGhouls Bhowling Alley Kiss of Death Wiked Cricket Grab Bag and More!!!brown_chocolate87643Nessuna valutazione finora
- Com Skills AnswersDocumento1 paginaCom Skills Answersbrown_chocolate87643Nessuna valutazione finora
- Communication Skills 11 ExaminationDocumento8 pagineCommunication Skills 11 Examinationbrown_chocolate87643Nessuna valutazione finora
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (121)
- Introduction JoviintroDocumento4 pagineIntroduction Joviintrobrown_chocolate87643Nessuna valutazione finora
- Harmonization of HR Policies - RoughDocumento119 pagineHarmonization of HR Policies - RoughFrancis SoiNessuna valutazione finora
- ESL Discussion Questions To Practise IdiomsDocumento5 pagineESL Discussion Questions To Practise IdiomsElaine Mc AuliffeNessuna valutazione finora
- D 5431 - 93 Rdu0mzetotmDocumento4 pagineD 5431 - 93 Rdu0mzetotmJuanNessuna valutazione finora
- Grade 8 MAPEH ReviewerDocumento4 pagineGrade 8 MAPEH ReviewerVictoria DelgadoNessuna valutazione finora
- Pakistan RailwayDocumento38 paginePakistan RailwayمحمودعليNessuna valutazione finora
- 4mb/01r-Que-2023 Jun Edexcel PastpaperDocumento24 pagine4mb/01r-Que-2023 Jun Edexcel Pastpaperaugust.shwanNessuna valutazione finora
- Morning Star Cafe Menu Revised 08-14-2019 On Line Menu 1Documento2 pagineMorning Star Cafe Menu Revised 08-14-2019 On Line Menu 1api-471935951Nessuna valutazione finora
- Esab Ok 55 (3.2 MM)Documento1 paginaEsab Ok 55 (3.2 MM)DHANEESH VELLILAPULLINessuna valutazione finora
- Envgeo D 17 00052 - R1Documento42 pagineEnvgeo D 17 00052 - R1Recep AkanNessuna valutazione finora
- Updated CBRN - Guideline - Final - Dec 16 - 2021Documento110 pagineUpdated CBRN - Guideline - Final - Dec 16 - 2021Mohammed AbdillahiNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- R0221120 - Salsabilla Cindy Pratama - Daily ActivityDocumento15 pagineR0221120 - Salsabilla Cindy Pratama - Daily Activitysalsabilla cindy pratamaNessuna valutazione finora
- Ficha Tecnica p501Documento4 pagineFicha Tecnica p501LizbethNessuna valutazione finora
- Mass, Weigth and Density IGCSEDocumento6 pagineMass, Weigth and Density IGCSEsapiniNessuna valutazione finora
- The Guardian Weekly 9-02-2024-2Documento64 pagineThe Guardian Weekly 9-02-2024-2krris.galNessuna valutazione finora
- Advent of SkyscrapersDocumento27 pagineAdvent of SkyscrapersJoshua AlboNessuna valutazione finora
- Key-Sentence Writing 1Documento23 pagineKey-Sentence Writing 1Thảo VyNessuna valutazione finora
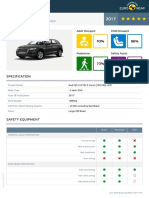
- Euroncap 2017 Audi q5 DatasheetDocumento13 pagineEuroncap 2017 Audi q5 DatasheetCosmin AnculiaNessuna valutazione finora
- Supplier S Documentation of Equipment PDFDocumento32 pagineSupplier S Documentation of Equipment PDFzhangjieNessuna valutazione finora
- Booster Mock Test For NEETDocumento15 pagineBooster Mock Test For NEETDrNessuna valutazione finora
- Bravo Petroleum ProfileDocumento8 pagineBravo Petroleum ProfileJacob Thom ZiwahwaNessuna valutazione finora
- 4g Frequency BandsDocumento5 pagine4g Frequency BandsaorukoNessuna valutazione finora
- How I Quit SmokingDocumento17 pagineHow I Quit Smokingquitsmoking100% (3)
- A. Desplat: Godzilla (2014) - Film Score AnalysisDocumento18 pagineA. Desplat: Godzilla (2014) - Film Score AnalysisR.PercacciNessuna valutazione finora
- Ffu 0000034 01Documento8 pagineFfu 0000034 01Karunia LestariNessuna valutazione finora
- ATF - List of Explosive MaterialsDocumento8 pagineATF - List of Explosive MaterialsxiaotaoscribdNessuna valutazione finora
- Rama Varma Anagha Research PaperDocumento12 pagineRama Varma Anagha Research Paperapi-308560676Nessuna valutazione finora
- JNTU Old Question Papers 2007Documento8 pagineJNTU Old Question Papers 2007Srinivasa Rao GNessuna valutazione finora
- Documentation ExamplesDocumento5 pagineDocumentation ExamplesErika HarveryNessuna valutazione finora
- Unit 1 - Theoretical Framework PDFDocumento67 pagineUnit 1 - Theoretical Framework PDFMeli EujaniánNessuna valutazione finora
- 06 Brakes PDFDocumento6 pagine06 Brakes PDFAKILI PSINessuna valutazione finora
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDDa EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDValutazione: 5 su 5 stelle5/5 (2)
- The Age of Magical Overthinking: Notes on Modern IrrationalityDa EverandThe Age of Magical Overthinking: Notes on Modern IrrationalityValutazione: 4 su 5 stelle4/5 (28)
- LIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionDa EverandLIT: Life Ignition Tools: Use Nature's Playbook to Energize Your Brain, Spark Ideas, and Ignite ActionValutazione: 4 su 5 stelle4/5 (404)
- Think This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeDa EverandThink This, Not That: 12 Mindshifts to Breakthrough Limiting Beliefs and Become Who You Were Born to BeValutazione: 2 su 5 stelle2/5 (1)
- Summary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedDa EverandSummary: The Psychology of Money: Timeless Lessons on Wealth, Greed, and Happiness by Morgan Housel: Key Takeaways, Summary & Analysis IncludedValutazione: 5 su 5 stelle5/5 (81)
- The Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaDa EverandThe Body Keeps the Score by Bessel Van der Kolk, M.D. - Book Summary: Brain, Mind, and Body in the Healing of TraumaValutazione: 4.5 su 5 stelle4.5/5 (266)
- Gut: the new and revised Sunday Times bestsellerDa EverandGut: the new and revised Sunday Times bestsellerValutazione: 4 su 5 stelle4/5 (393)