Potrebbero piacerti anche
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- Poster ExamplesDocumento6 paginePoster ExamplesJoel SanchezNessuna valutazione finora
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- CHE160A Syllabus S2014Documento6 pagineCHE160A Syllabus S2014Joel SanchezNessuna valutazione finora
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- Biosynthesis of NRPS PKS StructuresDocumento11 pagineBiosynthesis of NRPS PKS StructuresJoel SanchezNessuna valutazione finora
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- APpendix For Introduction To ThermodyanmicsDocumento2 pagineAPpendix For Introduction To ThermodyanmicsJoel SanchezNessuna valutazione finora
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Mocvd+Mbe Cee135 LectureDocumento20 pagineMocvd+Mbe Cee135 LectureJoel SanchezNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Chemical Engineering Lab ManualDocumento34 pagineChemical Engineering Lab ManualJoel SanchezNessuna valutazione finora
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- DLCP PV SOlarCellAppNoteDocumento16 pagineDLCP PV SOlarCellAppNoteVivek BelaNessuna valutazione finora
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Viscosity JsDocumento5 pagineViscosity JsJoel SanchezNessuna valutazione finora
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
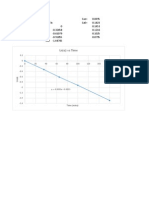
- Rate Constant Determination from Decay DataDocumento3 pagineRate Constant Determination from Decay DataJoel SanchezNessuna valutazione finora
- CH 01Documento102 pagineCH 01owned21cNessuna valutazione finora
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Solutions For Zemner Organic ChemistryDocumento6 pagineSolutions For Zemner Organic ChemistryJoel SanchezNessuna valutazione finora
- Biology 3-20 SecAss 12-1Documento1 paginaBiology 3-20 SecAss 12-1Edward Egan100% (1)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- ST - English 4 - Q3Documento19 pagineST - English 4 - Q3Joyce Mae OmerezNessuna valutazione finora
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- SCIENCE 10 (4TH Quarter) - Protein SynthesisDocumento4 pagineSCIENCE 10 (4TH Quarter) - Protein SynthesisJyña Khura TanoNessuna valutazione finora
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- Botanical Oils For Skin CareDocumento17 pagineBotanical Oils For Skin CareTrần NhỏNessuna valutazione finora
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Methods of Analysis: 5. Pharmaceutical Technical Procedures: 5.8 Methods of SterilizationDocumento5 pagineMethods of Analysis: 5. Pharmaceutical Technical Procedures: 5.8 Methods of Sterilizationrevy aprilliaNessuna valutazione finora
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Microfossils Description Paper: Palaeontology Practical: KalpanadekakalitaDocumento32 pagineMicrofossils Description Paper: Palaeontology Practical: Kalpanadekakalitarikalave ramanNessuna valutazione finora
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- Effects of Ethanol and Caffeine on Crayfish Heart RateDocumento1 paginaEffects of Ethanol and Caffeine on Crayfish Heart Ratebroddy1Nessuna valutazione finora
- Journal of Theoretical Biology: Qihua Huang, Hao Wang, Mark A. LewisDocumento19 pagineJournal of Theoretical Biology: Qihua Huang, Hao Wang, Mark A. Lewisyuukanishi6881Nessuna valutazione finora
- Cancer Cell Biology SyllabusDocumento2 pagineCancer Cell Biology SyllabusJose-Rico GonzalesNessuna valutazione finora
- Chapter 1 of Blue MindDocumento23 pagineChapter 1 of Blue MindSouthern California Public Radio100% (1)
- Soal Pat Inggris Kelas 7 - 2021Documento9 pagineSoal Pat Inggris Kelas 7 - 2021Bella Septiani FaryanNessuna valutazione finora
- Seroepidemiology of Clamaydia TDocumento8 pagineSeroepidemiology of Clamaydia TMulatuNessuna valutazione finora
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- 3rd Summative Test in English - Q1Documento4 pagine3rd Summative Test in English - Q1Gessle GamirNessuna valutazione finora
- TissuesDocumento23 pagineTissuesWubalem DestaNessuna valutazione finora
- Dr. Morse's Herbal Formulation ListDocumento10 pagineDr. Morse's Herbal Formulation ListTom50% (2)
- Sigma Services, Equipment, Instruments and Media Services overviewDocumento87 pagineSigma Services, Equipment, Instruments and Media Services overviewPreeti JaiswalNessuna valutazione finora
- TATIANADocumento8 pagineTATIANAKaizer NdoloNessuna valutazione finora
- Pneumonia Case Study: Zion FerrerDocumento40 paginePneumonia Case Study: Zion FerrerAllysa MacalinoNessuna valutazione finora
- Karakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiDocumento7 pagineKarakterisasi Koleksi Plasma Nutfah Tomat Lokal Dan IntroduksiAlexanderNessuna valutazione finora
- Systematic BotanyDocumento358 pagineSystematic Botanyrandel100% (3)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Journal of Theoretical Biology: Paolo Sibani, Simon ChristiansenDocumento5 pagineJournal of Theoretical Biology: Paolo Sibani, Simon ChristiansensdfklnNessuna valutazione finora
- Physiology - Last Minute Revision Points: DR - SreetejaDocumento25 paginePhysiology - Last Minute Revision Points: DR - SreetejaBanu KubendiranNessuna valutazione finora
- Water Indices For Water UsesDocumento12 pagineWater Indices For Water UsesWayaya2009Nessuna valutazione finora
- Fphar 12 741623Documento17 pagineFphar 12 741623Hendri AldratNessuna valutazione finora
- Human Anatomy Physiology Chapter 12 Sensory Organ NotesDocumento9 pagineHuman Anatomy Physiology Chapter 12 Sensory Organ NotesadarshNessuna valutazione finora
- Morphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesDocumento7 pagineMorphological Characterization of Pichia Guilliermondii and Saccharomyces Cerevisiae Yeast and Their Effects On Adherence of IntesAnonymous Y83Rx25Nessuna valutazione finora
- Methods, Method Verification and ValidationDocumento14 pagineMethods, Method Verification and Validationjljimenez1969100% (16)
- Lichens-Introduction, Classification, Structure, Reproduction and Economic Importance by Suresh Prashad Bhatta (M. Sc. Botany)Documento11 pagineLichens-Introduction, Classification, Structure, Reproduction and Economic Importance by Suresh Prashad Bhatta (M. Sc. Botany)Suresh BhattaNessuna valutazione finora
- EstomagoDocumento43 pagineEstomagoTanya FigueroaNessuna valutazione finora
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- Mbbs - Lecture Notes, Study Material and Important Questions, AnswersDocumento4 pagineMbbs - Lecture Notes, Study Material and Important Questions, AnswersM.V. TV67% (3)