Potrebbero piacerti anche
- Mariana Rodrigues RozattoDocumento101 pagineMariana Rodrigues RozattoCláudia Rachid ColussiNessuna valutazione finora
- 27-11-2007 16-16-07 Atividade Antioxidante de FlavonoidesDocumento8 pagine27-11-2007 16-16-07 Atividade Antioxidante de FlavonoidesDaniel De CarvalhoNessuna valutazione finora
- Antibiograma passo a passoDocumento29 pagineAntibiograma passo a passoVanessa SchmitzNessuna valutazione finora
- Infecção Nos Serviços de SaudeDocumento36 pagineInfecção Nos Serviços de SaudeCláudia Rachid ColussiNessuna valutazione finora
- Artigo CryptococcusDocumento12 pagineArtigo CryptococcusCláudia Rachid ColussiNessuna valutazione finora
- Diagnóstico laboratorial das dermatofitosesDocumento4 pagineDiagnóstico laboratorial das dermatofitosesDiogo Dias CordeiroNessuna valutazione finora
- Diagnóstico laboratorial das dermatofitosesDocumento4 pagineDiagnóstico laboratorial das dermatofitosesDiogo Dias CordeiroNessuna valutazione finora
- Microbiologia Laboratorial - Da Requisição de Exames A Análise MicrobiológicaDocumento28 pagineMicrobiologia Laboratorial - Da Requisição de Exames A Análise MicrobiológicaCláudia Rachid ColussiNessuna valutazione finora
- Artigo CryptococcusDocumento12 pagineArtigo CryptococcusCláudia Rachid ColussiNessuna valutazione finora
- Diagnóstico laboratorial das dermatofitosesDocumento4 pagineDiagnóstico laboratorial das dermatofitosesDiogo Dias CordeiroNessuna valutazione finora
- Apostilapratica 2 120911162509 Phpapp01Documento12 pagineApostilapratica 2 120911162509 Phpapp01Kelly BeliniNessuna valutazione finora
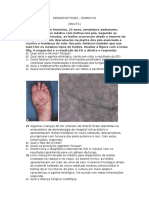
- DERMATOFITOSES - ExercícioDocumento2 pagineDERMATOFITOSES - ExercícioCláudia Rachid ColussiNessuna valutazione finora
- 01 MaláriaDocumento37 pagine01 MaláriaCláudia Rachid ColussiNessuna valutazione finora
- HEMOSTASIADocumento9 pagineHEMOSTASIACláudia Rachid Colussi100% (1)
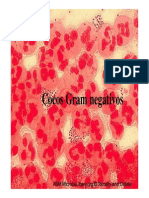
- Cocos Gram negativos causadores de infecçõesDocumento39 pagineCocos Gram negativos causadores de infecçõesCláudia Rachid ColussiNessuna valutazione finora
- 6+-+detecção+e+identificação+de+bactérias+de+importância+médicaDocumento150 pagine6+-+detecção+e+identificação+de+bactérias+de+importância+médicaThigo RossiNessuna valutazione finora
- Principais causas de variação em exames laboratoriaisDocumento55 paginePrincipais causas de variação em exames laboratoriaisCláudia Rachid ColussiNessuna valutazione finora
- Anemias HemolíticasDocumento30 pagineAnemias HemolíticasCláudia Rachid ColussiNessuna valutazione finora
- Endocrino Casos ClinicosDocumento20 pagineEndocrino Casos ClinicosCláudia Rachid ColussiNessuna valutazione finora
- ManualDeExames2013 HermesPardiniDocumento380 pagineManualDeExames2013 HermesPardiniLeiliane S. CardosoNessuna valutazione finora
- Classificacao de HelmintosDocumento4 pagineClassificacao de HelmintosCláudia Rachid ColussiNessuna valutazione finora
- HELMINTOSDocumento47 pagineHELMINTOSCláudia Rachid ColussiNessuna valutazione finora
- Introducao - HelmintologiaDocumento61 pagineIntroducao - HelmintologiaCláudia Rachid ColussiNessuna valutazione finora
- Classificacao de HelmintosDocumento4 pagineClassificacao de HelmintosCláudia Rachid ColussiNessuna valutazione finora
- Aulas Práticas de ParasitologiaDocumento67 pagineAulas Práticas de ParasitologiaCláudia Rachid ColussiNessuna valutazione finora
- Diagnóstico Bacteriológico Das Infecções Do Trato Urinário - Uma Revisão Técnica - ARTIGODocumento9 pagineDiagnóstico Bacteriológico Das Infecções Do Trato Urinário - Uma Revisão Técnica - ARTIGOCláudia Rachid ColussiNessuna valutazione finora
- Aulas Praticas de MicrobiologiaDocumento57 pagineAulas Praticas de MicrobiologiaNina Clara RochaNessuna valutazione finora
- Métodos Diagnósticos em ParasitologiaDocumento49 pagineMétodos Diagnósticos em ParasitologiaCláudia Rachid Colussi100% (1)
- Métodos Diagnósticos em ParasitologiaDocumento49 pagineMétodos Diagnósticos em ParasitologiaCláudia Rachid Colussi100% (1)
- XTZ 125 Xe 2014Documento136 pagineXTZ 125 Xe 2014Victor MirandaNessuna valutazione finora
- ResMédica - Aprovados e Classific - (Publicar)Documento246 pagineResMédica - Aprovados e Classific - (Publicar)Biih FialhoNessuna valutazione finora
- Voucher CintiaDocumento3 pagineVoucher CintiaPietra CrizolNessuna valutazione finora
- Acentuação de PalavrasDocumento44 pagineAcentuação de PalavrasEvandro RibeiroNessuna valutazione finora
- Resultado final QOCON Tec EAT/EIT 1-2019 avaliação curricular ADM São José dos CamposDocumento11 pagineResultado final QOCON Tec EAT/EIT 1-2019 avaliação curricular ADM São José dos CamposfidoruckNessuna valutazione finora
- Bhabha HomiDocumento10 pagineBhabha HomiPatrick BastosNessuna valutazione finora
- ATIV EXPL 14 - CPSN - Fundamentação TeóricaDocumento4 pagineATIV EXPL 14 - CPSN - Fundamentação Teóricadébora_christina_6Nessuna valutazione finora
- Cronograma de Estudos BNB Pos Edital Semanas 1 A 3Documento34 pagineCronograma de Estudos BNB Pos Edital Semanas 1 A 3joao marcosNessuna valutazione finora
- Exames de Matemática 12 Classe.Documento61 pagineExames de Matemática 12 Classe.Paulo Elias Sitoe0% (1)
- Análise de Cartas de ControleDocumento3 pagineAnálise de Cartas de ControleJúlia Stelvia GouveiaNessuna valutazione finora
- Análise de infográfico sobre desigualdades sociais no BrasilDocumento9 pagineAnálise de infográfico sobre desigualdades sociais no BrasilBel JamaicaNessuna valutazione finora
- FICHA TÉCNICA PILOTO NovaDocumento1 paginaFICHA TÉCNICA PILOTO NovaGADJNessuna valutazione finora
- Neuropsicologia do envelhecimentoDocumento37 pagineNeuropsicologia do envelhecimentoJoão Paulo MoraisNessuna valutazione finora
- NIE Dimel 13 - 09Documento6 pagineNIE Dimel 13 - 09Debora EmyNessuna valutazione finora
- CW 3-1Documento18 pagineCW 3-1guixcontactNessuna valutazione finora
- Trabalho Equações Do 2° Grau - AdaptadaDocumento2 pagineTrabalho Equações Do 2° Grau - AdaptadaGessica ByczkovskiNessuna valutazione finora
- Cooperação com DeusDocumento65 pagineCooperação com DeusFatima4BretzNessuna valutazione finora
- Responsabilidade Social Das EmpresasDocumento62 pagineResponsabilidade Social Das EmpresasNhaueleque Júnior100% (7)
- 11º Ano - Teste Geografia ErtDocumento5 pagine11º Ano - Teste Geografia ErtTobiasKnight100% (2)
- Lista de Exercícios AV1.Documento4 pagineLista de Exercícios AV1.Carlos BrandãoNessuna valutazione finora
- RAEYMAEKER, Luís de Filosofia Do Ser São Paulo, Editora Herder, 1967Documento173 pagineRAEYMAEKER, Luís de Filosofia Do Ser São Paulo, Editora Herder, 1967rvfortes100% (2)
- Planejamento Maio A1 - B1Documento4 paginePlanejamento Maio A1 - B1Francisca AlencarNessuna valutazione finora
- Isomeria geométrica e óptica no ibuprofenoDocumento7 pagineIsomeria geométrica e óptica no ibuprofenoRhuan MendesNessuna valutazione finora
- RELATÓRIO DE PROJETO FUNDIÇÃO - FinalDocumento13 pagineRELATÓRIO DE PROJETO FUNDIÇÃO - FinalVinicius CamposNessuna valutazione finora
- Laboratório de Física Geral 3Documento78 pagineLaboratório de Física Geral 3José Lucas Decote de Carvalho LirioNessuna valutazione finora
- Breve Introdução A Mecânica de LagrangeDocumento4 pagineBreve Introdução A Mecânica de Lagrangeantonio ricardo 010% (1)
- SKF PER.5203RYY2 SpecificationDocumento2 pagineSKF PER.5203RYY2 SpecificationRanielNessuna valutazione finora
- DIRETRIZ CNBC 01-2012 Codigo Ética Bombeiro Civil e Pessoal de Serviços de Emergências - 4a EdDocumento15 pagineDIRETRIZ CNBC 01-2012 Codigo Ética Bombeiro Civil e Pessoal de Serviços de Emergências - 4a Edivan pesquisaNessuna valutazione finora
- Engenharia Civil fluxogramaDocumento8 pagineEngenharia Civil fluxogramaElitiere Silva CruzNessuna valutazione finora