Potrebbero piacerti anche
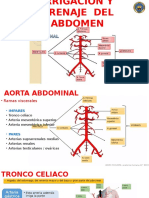
- Vascular AbdomenDocumento21 pagineVascular AbdomenStalynNñz100% (6)
- Informática SEM-Informe S9-Collantes-GRUPO24Documento7 pagineInformática SEM-Informe S9-Collantes-GRUPO24franco mamani rodriguezNessuna valutazione finora
- Biomecánica y Biodinámica EXPO COMPLETODocumento16 pagineBiomecánica y Biodinámica EXPO COMPLETOJosé mario Vázquez poloNessuna valutazione finora
- Seminario Fisica Semana 13 BIOELECTRICIDADDocumento16 pagineSeminario Fisica Semana 13 BIOELECTRICIDADJoaquin Alcantara De la TorreNessuna valutazione finora
- 10 InformeDocumento4 pagine10 InformeECO- SANDNessuna valutazione finora
- Listado de BioquimicosDocumento126 pagineListado de BioquimicosJean Ezekielo De KentNessuna valutazione finora
- Examen Sistema Nervioso Grado Octavo Iete 2017Documento4 pagineExamen Sistema Nervioso Grado Octavo Iete 2017Diana Carolina Burbano Ruano80% (5)
- Necesidad de Eliminacion IntestinalDocumento15 pagineNecesidad de Eliminacion IntestinalJacob Bell100% (20)
- PericitosDocumento8 paginePericitosMaría Victoria BedoyaNessuna valutazione finora
- Práctica de Física - DensidadDocumento16 paginePráctica de Física - DensidadMonica VillalobosNessuna valutazione finora
- Pe RicitosDocumento3 paginePe RicitosHernán Rodriguez OtinianoNessuna valutazione finora
- Informe 10 - Grupo 26CDocumento7 pagineInforme 10 - Grupo 26CJenny TarrilloNessuna valutazione finora
- Informe s6 - Embrio Práctica - Grupo 18Documento8 pagineInforme s6 - Embrio Práctica - Grupo 18Shirleyxitha Carrero CastroNessuna valutazione finora
- Seminario 14 RADIOISOTOPOSDocumento18 pagineSeminario 14 RADIOISOTOPOShavr91Nessuna valutazione finora
- Bio Sem S9Documento4 pagineBio Sem S9Xiomara EvangelistaNessuna valutazione finora
- Tarea 6 FisicaDocumento6 pagineTarea 6 FisicaCeleste VásquezNessuna valutazione finora
- Informe 11 Embriología PrácticaDocumento9 pagineInforme 11 Embriología PrácticaEmery Sofía C. D. BustamanteNessuna valutazione finora
- CoriDocumento4 pagineCoriKenner Poult Santisteban MuñozNessuna valutazione finora
- Informe Anatomía Humana II Sesión 3Documento7 pagineInforme Anatomía Humana II Sesión 3Angie Flores MamaniNessuna valutazione finora
- Inmunologia en El EmbarazoDocumento36 pagineInmunologia en El EmbarazoDioser AvilaNessuna valutazione finora
- Informe 5 Embriología SeminarioDocumento7 pagineInforme 5 Embriología SeminarioEmery Sofía C. D. BustamanteNessuna valutazione finora
- BioelectricidadDocumento32 pagineBioelectricidadMaritho Vera SànchezNessuna valutazione finora
- Lectura 01 - Membrana de Transporte y Transporte Vesicular - En.esDocumento6 pagineLectura 01 - Membrana de Transporte y Transporte Vesicular - En.esNely HuertaNessuna valutazione finora
- Informe S8 Psicología SeminarioDocumento7 pagineInforme S8 Psicología SeminarioEmery Sofía C. D. BustamanteNessuna valutazione finora
- Tejido conectivo-2020-USMP PDFDocumento54 pagineTejido conectivo-2020-USMP PDFSergio Contreras RomeroNessuna valutazione finora
- Placenta MacroscópicaDocumento11 paginePlacenta MacroscópicaLiliana Julissa García AlarcónNessuna valutazione finora
- Informe S10 PBM PrácticaDocumento13 pagineInforme S10 PBM PrácticaAlexander E. Bruckmann AvilaNessuna valutazione finora
- Informe S5-SP PracticaDocumento6 pagineInforme S5-SP PracticajuanNessuna valutazione finora
- Informe S4 Psico Sem - Grupo 23Documento4 pagineInforme S4 Psico Sem - Grupo 23Emery Sofía C. D. BustamanteNessuna valutazione finora
- HemocomDocumento17 pagineHemocomguillermoNessuna valutazione finora
- Salud Pública - SEMINARIO - Informe 2Documento5 pagineSalud Pública - SEMINARIO - Informe 2Xiomara Janne Linares SaldañaNessuna valutazione finora
- Informe 05 - Inmunología Sem - Grupo 19Documento13 pagineInforme 05 - Inmunología Sem - Grupo 19Jenny TarrilloNessuna valutazione finora
- Embriología PRAC - EXPON°11-OBSERVACION DE EMBRIONESDocumento19 pagineEmbriología PRAC - EXPON°11-OBSERVACION DE EMBRIONESMiguelIvanB.GuerreroNessuna valutazione finora
- BIOELECTRICIDADDocumento3 pagineBIOELECTRICIDADChristian Raul Rojas TarrilloNessuna valutazione finora
- Informe 5.7Documento9 pagineInforme 5.7Cielo Suarez VeasNessuna valutazione finora
- BIOESTADITICA - S4 - InformeDocumento11 pagineBIOESTADITICA - S4 - InformeNayeli Castillo MinianoNessuna valutazione finora
- InformeDocumento6 pagineInformeGustavo Herrera MontenegroNessuna valutazione finora
- INFORME #09 - GRUPO 36 - BIOELECTRICIDAD - DoxxDocumento8 pagineINFORME #09 - GRUPO 36 - BIOELECTRICIDAD - DoxxJosé mario Vázquez poloNessuna valutazione finora
- Informe 3 Embriología SeminarioDocumento8 pagineInforme 3 Embriología SeminarioEmery Sofía C. D. BustamanteNessuna valutazione finora
- Informe S12 Psico Sem - Grupo 23Documento6 pagineInforme S12 Psico Sem - Grupo 23Emery Sofía C. D. BustamanteNessuna valutazione finora
- Tórax Parietal, Aparato Respiratorio y Aparato CardiovascularDocumento17 pagineTórax Parietal, Aparato Respiratorio y Aparato CardiovascularElianne Alexandra Diaz SaldañaNessuna valutazione finora
- Informe S11 Psico SemDocumento9 pagineInforme S11 Psico SemEmery Sofía C. D. BustamanteNessuna valutazione finora
- Mitocondrias y ADN Mitocondrial.Documento24 pagineMitocondrias y ADN Mitocondrial.Daniel Clavijo100% (1)
- Informe S10 Psico SemDocumento7 pagineInforme S10 Psico SemEmery Sofía C. D. BustamanteNessuna valutazione finora
- Pregunta 2 Informe PFDocumento2 paginePregunta 2 Informe PFAna Paula FernandezNessuna valutazione finora
- Informe 12 - Informatica IDocumento9 pagineInforme 12 - Informatica IHector Arbañil NanfuñayNessuna valutazione finora
- Informe 12 Embriología PrácticaDocumento7 pagineInforme 12 Embriología PrácticaEmery Sofía C. D. BustamanteNessuna valutazione finora
- Hidrostática I y IIDocumento5 pagineHidrostática I y IICelesteNessuna valutazione finora
- INFORME S11 Informática IIDocumento8 pagineINFORME S11 Informática IIXiomara Janne Linares SaldañaNessuna valutazione finora
- Citoplazma, Citoesqueleto y Matriz ExtracelularDocumento10 pagineCitoplazma, Citoesqueleto y Matriz ExtracelularSofia Arias-StellaNessuna valutazione finora
- Trab Preg 3 Sem BioDocumento13 pagineTrab Preg 3 Sem BioMargaritaty0% (1)
- Seminario N 5Documento16 pagineSeminario N 5Nina Saboya CórdovaNessuna valutazione finora
- Informe 3 Histología PrácticaDocumento9 pagineInforme 3 Histología PrácticaEmery Sofía C. D. BustamanteNessuna valutazione finora
- Transporte IntracelularDocumento7 pagineTransporte IntracelularRamiro Santamaria100% (1)
- Exposición Semana 10 (Grupo 10T 25B)Documento14 pagineExposición Semana 10 (Grupo 10T 25B)Antenor Rodas Sembrera100% (1)
- Taller #1. ESTRUCTURA Y FUNCIÓN DE LA CÉLULADocumento2 pagineTaller #1. ESTRUCTURA Y FUNCIÓN DE LA CÉLULAJuliana PerdomoNessuna valutazione finora
- El Origen de La Vida Terrestre ModernaDocumento6 pagineEl Origen de La Vida Terrestre ModernaEnrique Moncada MapelliNessuna valutazione finora
- Informe 12 Histología SeminarioDocumento10 pagineInforme 12 Histología SeminarioEmery Sofía C. D. BustamanteNessuna valutazione finora
- Tarea 1 Histología 03T22Documento13 pagineTarea 1 Histología 03T22Jenny TarrilloNessuna valutazione finora
- Observacion de Movimiento CelularDocumento19 pagineObservacion de Movimiento CelularAlex Hinostroza Huaman0% (1)
- INFORME S12 - GRUPO Jquirozv-01Documento11 pagineINFORME S12 - GRUPO Jquirozv-01Sadae BDNessuna valutazione finora
- Cell MadresDocumento16 pagineCell MadresEdmundo CisnerosNessuna valutazione finora
- Reprogramación NuclearDocumento52 pagineReprogramación NuclearAlfonso MédgerNessuna valutazione finora
- Celulas TroncalesDocumento6 pagineCelulas TroncalesTanya Fuentealba NegrónNessuna valutazione finora
- Capitulo 20 Virosis - 1Documento10 pagineCapitulo 20 Virosis - 1Carlos PeredaNessuna valutazione finora
- Neuropatía DiabéticaDocumento4 pagineNeuropatía DiabéticaCarlos PeredaNessuna valutazione finora
- Capitulo 3 - MaculosasDocumento8 pagineCapitulo 3 - MaculosasCarlos PeredaNessuna valutazione finora
- Capitulo 2 - Lesiones ElementalesDocumento5 pagineCapitulo 2 - Lesiones ElementalesCarlos PeredaNessuna valutazione finora
- Capitulo 15 - Eritema Multiforme 1Documento9 pagineCapitulo 15 - Eritema Multiforme 1Carlos PeredaNessuna valutazione finora
- Actualidad Cardiologia Clinica Mayo 2019Documento8 pagineActualidad Cardiologia Clinica Mayo 2019Carlos PeredaNessuna valutazione finora
- Repetto - Teorico. Meningitis Bacteriana 2019Documento39 pagineRepetto - Teorico. Meningitis Bacteriana 2019Carlos Pereda100% (1)
- Hormona Antimulleriana Por. Mónica GuizburgDocumento6 pagineHormona Antimulleriana Por. Mónica GuizburgCarlos PeredaNessuna valutazione finora
- Parcial 1 de Micro 2 Catedra 1 25Documento5 pagineParcial 1 de Micro 2 Catedra 1 25Carlos PeredaNessuna valutazione finora
- Folleto Cobas 8000 - 2013Documento4 pagineFolleto Cobas 8000 - 2013Carlos PeredaNessuna valutazione finora
- Hormona Antimulleriana Por. Mónica GuizburgDocumento6 pagineHormona Antimulleriana Por. Mónica GuizburgCarlos PeredaNessuna valutazione finora
- E MergenciaDocumento13 pagineE MergenciaCarlos PeredaNessuna valutazione finora
- Folleto Cobas 8000 - 2013 PDFDocumento16 pagineFolleto Cobas 8000 - 2013 PDFCarlos Pereda100% (1)
- AmuchasteguiDocumento3 pagineAmuchasteguiCarlos PeredaNessuna valutazione finora
- Hipnosis PDFDocumento9 pagineHipnosis PDFCarlos PeredaNessuna valutazione finora
- El Surgimiento de La Complementariedad Niels Bohr y La Conferencia de ComoDocumento12 pagineEl Surgimiento de La Complementariedad Niels Bohr y La Conferencia de ComoCarlos PeredaNessuna valutazione finora
- Tej LinfoideDocumento6 pagineTej LinfoideCarlos PeredaNessuna valutazione finora
- Evans Gambit 5... Ae7 - ChessForReal - Com &Documento39 pagineEvans Gambit 5... Ae7 - ChessForReal - Com &supramagusNessuna valutazione finora
- Bibliografia Normas de VancouverDocumento6 pagineBibliografia Normas de VancouverFco SegoviaNessuna valutazione finora
- Protein AsDocumento14 pagineProtein AsCarlos PeredaNessuna valutazione finora
- 220 DerivadasParcialesSegundasPolinomiosTaylorDocumento11 pagine220 DerivadasParcialesSegundasPolinomiosTaylorelsupertinchoNessuna valutazione finora
- Etica ProfesionalDocumento8 pagineEtica ProfesionalCarlos PeredaNessuna valutazione finora
- Reg LinealDocumento11 pagineReg LinealCarlos PeredaNessuna valutazione finora
- BrownieDocumento6 pagineBrownieCarlos PeredaNessuna valutazione finora
- 35 1 09Documento6 pagine35 1 09Carlos PeredaNessuna valutazione finora
- EcdisomaDocumento3 pagineEcdisomaCarlos PeredaNessuna valutazione finora
- XilenoDocumento4 pagineXilenoCarlos PeredaNessuna valutazione finora
- Levotiroxina Administrada en La Mañana VS NocheDocumento62 pagineLevotiroxina Administrada en La Mañana VS NocheJonathan Timmy Tornner YupanguiNessuna valutazione finora
- Teoria de Los 5 ElementosDocumento18 pagineTeoria de Los 5 Elementosmsomaza86% (22)
- Resumen Embriología Formación de La CaraDocumento5 pagineResumen Embriología Formación de La CaraMiguel AngelNessuna valutazione finora
- Wuolah Free 15Documento27 pagineWuolah Free 15Blanca Pérez SanzNessuna valutazione finora
- Aparato ReproductorDocumento3 pagineAparato ReproductorChiki JerezNessuna valutazione finora
- Seminario Incompatibilidad Feto-Materno Abo RHDocumento24 pagineSeminario Incompatibilidad Feto-Materno Abo RHjeanNessuna valutazione finora
- Radiologia RenalDocumento34 pagineRadiologia RenalJeniree OmañaNessuna valutazione finora
- RETICULOCITODocumento6 pagineRETICULOCITOMaribel Quispe CondoriNessuna valutazione finora
- Anatomía y Fisiología Del Aparato ReproductorDocumento9 pagineAnatomía y Fisiología Del Aparato ReproductorMaria Estela Mercado MartinezNessuna valutazione finora
- Fisiología Veterinaria - (Tema 64)Documento12 pagineFisiología Veterinaria - (Tema 64)JesusPelaezz100% (1)
- MOTIVACION Es La Propiedad Que Nos Impulsa y Capacita A Ejecutar Una ActividadDocumento8 pagineMOTIVACION Es La Propiedad Que Nos Impulsa y Capacita A Ejecutar Una ActividadLalyNessuna valutazione finora
- Sindrome NefriticoDocumento19 pagineSindrome NefriticoPedro Jose Baquero MarinNessuna valutazione finora
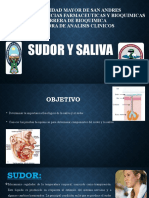
- Sudor y SalivaDocumento12 pagineSudor y SalivaAlejandra Fanola SalazarNessuna valutazione finora
- Fracturas Dentoalveolares 2Documento12 pagineFracturas Dentoalveolares 2Angely Emily Pérez DavilaNessuna valutazione finora
- Manual de Analisis de Modelos de OrtodonciaDocumento67 pagineManual de Analisis de Modelos de Ortodonciataniaqc25100% (2)
- 34 - 40 - MédulaDocumento7 pagine34 - 40 - MédulaTomas RodríguezNessuna valutazione finora
- Practica de FluidosDocumento2 paginePractica de FluidosTary0% (1)
- FISIOLOGIA I GB GR PDFDocumento12 pagineFISIOLOGIA I GB GR PDFHamletNessuna valutazione finora
- Vejiga y UretraDocumento3 pagineVejiga y UretraAcosta Karina 7 9 2 0 9 / 7Nessuna valutazione finora
- Efectos de Prostaglandinas y TromboxanosDocumento21 pagineEfectos de Prostaglandinas y TromboxanosYatziriNessuna valutazione finora
- El Agua en El Organismo AnimalDocumento53 pagineEl Agua en El Organismo Animalnataly_182100% (10)
- Recoleccion de CPH en NiñosDocumento4 pagineRecoleccion de CPH en NiñosERICK FERNANDO HUAINA CENIZARIONessuna valutazione finora
- Preguntas Expo PeriodonciaDocumento3 paginePreguntas Expo PeriodonciaJan's JNNessuna valutazione finora
- Esqueleto de La Cabeza Huesos de La Cintura Escapular: Anatomía y Fisiología HumanaDocumento7 pagineEsqueleto de La Cabeza Huesos de La Cintura Escapular: Anatomía y Fisiología HumanaDany DownieNessuna valutazione finora
- Medula Espinal Anatomia Funcion EstructuraDocumento9 pagineMedula Espinal Anatomia Funcion EstructuraRaquel Villavicencio BautistaNessuna valutazione finora
- 1 - Introduccíon A La AHDocumento24 pagine1 - Introduccíon A La AHErlin VargasNessuna valutazione finora
- Vesicual Biliar y RadiologiaDocumento21 pagineVesicual Biliar y RadiologiaELIZABETH MONDRAGON DIAZNessuna valutazione finora