Potrebbero piacerti anche
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (894)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Mirnomics: Microrna Biology and Computational AnalysisDocumento336 pagineMirnomics: Microrna Biology and Computational AnalysisJavierElectricoNessuna valutazione finora
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- Reviewer - Immunoserology - Part 2 PDFDocumento33 pagineReviewer - Immunoserology - Part 2 PDFJoshua TrinidadNessuna valutazione finora
- Bio Sol SheetDocumento6 pagineBio Sol Sheetapi-110789702Nessuna valutazione finora
- The Impact of Tumor Biology On Cancer Treatment and Multidisciplinary Strategies - M. Molls, Et Al., (Springer, 2009) WWDocumento363 pagineThe Impact of Tumor Biology On Cancer Treatment and Multidisciplinary Strategies - M. Molls, Et Al., (Springer, 2009) WWiuliNessuna valutazione finora
- Pathophysiology Test Bank Ch. 2-4Documento26 paginePathophysiology Test Bank Ch. 2-4Joyy100% (3)
- Studi Variasi Morfologi Dan Pola Pita Isozim Lundi Putih (Coleoptera: Melolonthidae) Yang Berperan Sebagai Hama Tanaman Salak Di Lereng Gunung MerapiDocumento5 pagineStudi Variasi Morfologi Dan Pola Pita Isozim Lundi Putih (Coleoptera: Melolonthidae) Yang Berperan Sebagai Hama Tanaman Salak Di Lereng Gunung MerapiSiti Nur AzizahNessuna valutazione finora
- TumbDocumento13 pagineTumbSiti Nur AzizahNessuna valutazione finora
- 2403 1 4801 1 10 20160210Documento6 pagine2403 1 4801 1 10 20160210eniNessuna valutazione finora
- Chelex PDFDocumento3 pagineChelex PDFSiti Nur AzizahNessuna valutazione finora
- Tugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Documento4 pagineTugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Siti Nur AzizahNessuna valutazione finora
- SujarwatiDocumento5 pagineSujarwatiSiti Nur AzizahNessuna valutazione finora
- Fulltext 9854Documento173 pagineFulltext 9854Siti Nur AzizahNessuna valutazione finora
- Bakteriosin PDFDocumento13 pagineBakteriosin PDFSiti Nur AzizahNessuna valutazione finora
- PTTI 2015 Unsoed 31okt14Documento3 paginePTTI 2015 Unsoed 31okt14Siti Nur AzizahNessuna valutazione finora
- 13 GurguDocumento8 pagine13 GurguSiti Nur AzizahNessuna valutazione finora
- Ektoparasit InggrisDocumento9 pagineEktoparasit InggrisSiti Nur AzizahNessuna valutazione finora
- Commission On Intellectual Property RightsDocumento107 pagineCommission On Intellectual Property RightsSiti Nur AzizahNessuna valutazione finora
- Lampiran Fix 22Documento9 pagineLampiran Fix 22Siti Nur AzizahNessuna valutazione finora
- Tugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Documento4 pagineTugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Siti Nur AzizahNessuna valutazione finora
- SujarwatiDocumento5 pagineSujarwatiSiti Nur AzizahNessuna valutazione finora
- Tentang IPRDocumento117 pagineTentang IPRSiti Nur AzizahNessuna valutazione finora
- My Daily Activity: Name: Siti Nur Azizah NIM: B1J011086Documento1 paginaMy Daily Activity: Name: Siti Nur Azizah NIM: B1J011086Siti Nur AzizahNessuna valutazione finora
- Bilal Mizra Verspagen 2006Documento10 pagineBilal Mizra Verspagen 2006Siti Nur AzizahNessuna valutazione finora
- Tugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Documento4 pagineTugas Kuliah Virologi Kelas B Siti Nur Azizah B1J011086Siti Nur AzizahNessuna valutazione finora
- Commission On Intellectual Property RightsDocumento107 pagineCommission On Intellectual Property RightsSiti Nur AzizahNessuna valutazione finora
- Commission On Intellectual Property RightsDocumento107 pagineCommission On Intellectual Property RightsSiti Nur AzizahNessuna valutazione finora
- Simple Present TenseDocumento3 pagineSimple Present TenseSiti Nur AzizahNessuna valutazione finora
- Jurnal Inggris Acara 1Documento9 pagineJurnal Inggris Acara 1Siti Nur AzizahNessuna valutazione finora
- Responses of Midbrain Auditory Neurons inDocumento7 pagineResponses of Midbrain Auditory Neurons inSiti Nur AzizahNessuna valutazione finora
- TaksheDocumento13 pagineTaksheSiti Nur AzizahNessuna valutazione finora
- Jurnal Fishe Nutrisi (Yoke Astriani, Siti Nur Azizah, Korrie Salsabila)Documento16 pagineJurnal Fishe Nutrisi (Yoke Astriani, Siti Nur Azizah, Korrie Salsabila)Siti Nur AzizahNessuna valutazione finora
- Heat Shock ProteinsDocumento4 pagineHeat Shock ProteinsAndri Praja SatriaNessuna valutazione finora
- Enzymes PresDocumento34 pagineEnzymes Presm. uma deviNessuna valutazione finora
- Liver Function Reviewer Cc2Documento7 pagineLiver Function Reviewer Cc2Leo NoquilNessuna valutazione finora
- SIYENSIKULADocumento3 pagineSIYENSIKULANoob KidNessuna valutazione finora
- Uric AcidDocumento16 pagineUric AcidMustafa KhandgawiNessuna valutazione finora
- RAPD-PCR Based Marker Approach For The Genetic Differentiation of Two Species of Cockroach (Order-Dictyoptera)Documento6 pagineRAPD-PCR Based Marker Approach For The Genetic Differentiation of Two Species of Cockroach (Order-Dictyoptera)LifedavidNessuna valutazione finora
- 1 s2.0 S2468124523000086 MainDocumento16 pagine1 s2.0 S2468124523000086 MainDana MuraneviciNessuna valutazione finora
- 2012 JuneDocumento198 pagine2012 JunealiceNessuna valutazione finora
- Classes of Antibiotics PDFDocumento1 paginaClasses of Antibiotics PDFVương TúNessuna valutazione finora
- The Biological Revolution: Periodicals of Engineering and Natural Sciences Vol. 3 No. 1 (2015) Available Online atDocumento2 pagineThe Biological Revolution: Periodicals of Engineering and Natural Sciences Vol. 3 No. 1 (2015) Available Online atAngel TaylorNessuna valutazione finora
- General Pharmacology-QuestionDocumento4 pagineGeneral Pharmacology-Questionlina amjadNessuna valutazione finora
- Section A Multiple Choice QuestionsDocumento6 pagineSection A Multiple Choice QuestionsViola Voon Li WeiNessuna valutazione finora
- Modern and Convensional Wound Dressing To Interleukin 1 and Interleukin 6 in Diabetic WoundDocumento2 pagineModern and Convensional Wound Dressing To Interleukin 1 and Interleukin 6 in Diabetic WoundGilang yuanggaNessuna valutazione finora
- 2&3 - Action Potential and Resting MembraneDocumento28 pagine2&3 - Action Potential and Resting MembraneTejesswinNessuna valutazione finora
- CELL CYCLE PresentationDocumento22 pagineCELL CYCLE PresentationCalvin YusopNessuna valutazione finora
- Cell organelles and their functionsDocumento1 paginaCell organelles and their functionsNazimNessuna valutazione finora
- Gen Physiology 2015Documento423 pagineGen Physiology 2015cathlynjoy.marsamoloNessuna valutazione finora
- A General Biology I Q1M1.2Documento26 pagineA General Biology I Q1M1.2Queng Eledia0% (1)
- Genetics of Axis Specification in DrosophilaDocumento3 pagineGenetics of Axis Specification in DrosophilaJohnkevin NocheNessuna valutazione finora
- Hemoglobin SigmaDocumento3 pagineHemoglobin SigmascheekoNessuna valutazione finora
- Beta Oxidation of Fatty AcidsDocumento4 pagineBeta Oxidation of Fatty AcidsJonas Galeos0% (1)
- Expression CloningDocumento16 pagineExpression CloningabrahamNessuna valutazione finora
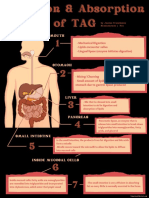
- Digestion & Absorption of TAGDocumento1 paginaDigestion & Absorption of TAGJanine Franchesca SuministradoNessuna valutazione finora
- CAPE Biology Factors Limiting PhotosynthesisDocumento2 pagineCAPE Biology Factors Limiting PhotosynthesisKashim AliNessuna valutazione finora
- IFC Product - U.S. Feather Meal-20150420074556Documento1 paginaIFC Product - U.S. Feather Meal-20150420074556Dinda AyuNessuna valutazione finora