Potrebbero piacerti anche
- BARRIENTOS - Alteraciones en La Integridad Del Acrosoma y de La Teca Perinuclear en Semen Criopreservado de VerracoDocumento8 pagineBARRIENTOS - Alteraciones en La Integridad Del Acrosoma y de La Teca Perinuclear en Semen Criopreservado de VerracoJJHTSCRIBDNessuna valutazione finora
- La Criopreservación Del Semen Porcino - Albéitar Portal VeterinariaDocumento3 pagineLa Criopreservación Del Semen Porcino - Albéitar Portal VeterinariaJJHTSCRIBDNessuna valutazione finora
- La Criopreservación Con Α-Tocoferol y La Selección Con Sephadex Mejoran La Calidad Del Semen PorcinoDocumento2 pagineLa Criopreservación Con Α-Tocoferol y La Selección Con Sephadex Mejoran La Calidad Del Semen PorcinoJJHTSCRIBDNessuna valutazione finora
- Ia PorcinoDocumento64 pagineIa PorcinoJJHTSCRIBD100% (1)
- Efecto de La Concentración Espermática de Semen Porcino Sobre Sus Características Post-DescongelamientoDocumento36 pagineEfecto de La Concentración Espermática de Semen Porcino Sobre Sus Características Post-DescongelamientoJJHTSCRIBDNessuna valutazione finora
- Efecto de Dos Métodos de Congelación Sobre La Viabilidad Espermática de Semen de VerracoDocumento5 pagineEfecto de Dos Métodos de Congelación Sobre La Viabilidad Espermática de Semen de VerracoJJHTSCRIBDNessuna valutazione finora
- WiggersDocumento4 pagineWiggersDani HdzNessuna valutazione finora
- Criopreservacion de Semen Suino en VenezuelaDocumento12 pagineCriopreservacion de Semen Suino en VenezuelaJJHTSCRIBDNessuna valutazione finora
- Fases Previa y Postcongelación Del Semen de Verraco en Pajillas de 5 ML y Capacidad de Fecundación de Los EspermatozoidesDocumento8 pagineFases Previa y Postcongelación Del Semen de Verraco en Pajillas de 5 ML y Capacidad de Fecundación de Los EspermatozoidesJJHTSCRIBDNessuna valutazione finora
- Flujo MultifásicoDocumento42 pagineFlujo MultifásicoAnna SanchezNessuna valutazione finora
- Intereses Del Conocimiento HabermasDocumento16 pagineIntereses Del Conocimiento Habermasjohnfcg1980Nessuna valutazione finora
- Purinas y Pirimidinas 2010Documento81 paginePurinas y Pirimidinas 2010joe_fox2132296Nessuna valutazione finora
- Respuestas Fisiológicas Al CalorDocumento62 pagineRespuestas Fisiológicas Al Calorpajarillo0Nessuna valutazione finora
- TCP 1Documento12 pagineTCP 1Joseph A. VargasNessuna valutazione finora
- Aprendizaje CognitivoDocumento12 pagineAprendizaje CognitivoEduardo Bravo LangeNessuna valutazione finora
- Papel Bioetica en La SociedadDocumento4 paginePapel Bioetica en La SociedadJJHTSCRIBDNessuna valutazione finora
- Qué Es Un Mapa ConceptualDocumento6 pagineQué Es Un Mapa ConceptualKaren RojazNessuna valutazione finora
- Auto Estim ADocumento4 pagineAuto Estim AJJHTSCRIBDNessuna valutazione finora
- TEST VAC Por Jorge Neira SilvaDocumento6 pagineTEST VAC Por Jorge Neira Silvamasterload100% (2)
- El Desarrollo de La Autoestima en AdolescentesDocumento4 pagineEl Desarrollo de La Autoestima en Adolescentesmonicadiazbesada100% (2)
- CiclocelularmitosisDocumento5 pagineCiclocelularmitosisWilliamNessuna valutazione finora
- Dirofilariosis Cardiopulmonar CaninaDocumento10 pagineDirofilariosis Cardiopulmonar CaninaJJHTSCRIBDNessuna valutazione finora
- ESKARETHDocumento29 pagineESKARETHRafael DomNessuna valutazione finora
- Anatomía HipofisisDocumento3 pagineAnatomía Hipofisispatymend23Nessuna valutazione finora
- Libro New NewDocumento611 pagineLibro New NewMax Lewiss Mafaldo DavilaNessuna valutazione finora
- La Vida en El Agua PDFDocumento12 pagineLa Vida en El Agua PDFEri-k GuzToNessuna valutazione finora
- Monografia ForenseDocumento12 pagineMonografia ForenseGloryIncaFuentes100% (1)
- MORRIS Psicologia Cap2Documento28 pagineMORRIS Psicologia Cap2Etpsi Trabajo Psicologico100% (2)
- CESTODOSDocumento4 pagineCESTODOSPalooma KnightNessuna valutazione finora
- Cupcakes para PerrosDocumento13 pagineCupcakes para PerrosAna López GarcíaNessuna valutazione finora
- Introducción Del Curso Entomología y MIPDocumento13 pagineIntroducción Del Curso Entomología y MIPAndres David PadillaNessuna valutazione finora
- La VacaDocumento5 pagineLa VacaCarmen Julia Sanchez BustamanteNessuna valutazione finora
- Orden OrthopteraDocumento12 pagineOrden OrthopteraJorge Ruiz67% (3)
- Parasitológico Simple y SeriadoDocumento16 pagineParasitológico Simple y SeriadoCarla Yelixa Garnica Meneces50% (6)
- Clase 1 Anemias Carenciales - Deficiencia de HierroDocumento45 pagineClase 1 Anemias Carenciales - Deficiencia de HierroedwinNessuna valutazione finora
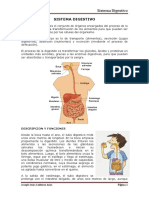
- Asignacion Sistema DigestivoDocumento6 pagineAsignacion Sistema DigestivoRichard Q.Nessuna valutazione finora
- Programa de Mejora de La Condición Física 3º EsoDocumento5 paginePrograma de Mejora de La Condición Física 3º Esotrivas60Nessuna valutazione finora
- Embriologia de Cara y OdontogenesisDocumento70 pagineEmbriologia de Cara y OdontogenesisAle Cordoba HerreraNessuna valutazione finora
- Clasififcacion AODocumento64 pagineClasififcacion AOLuisJorgeNessuna valutazione finora
- OLIGOAMNIOSDocumento16 pagineOLIGOAMNIOSKathy Santa Cruz ObesoNessuna valutazione finora
- Clase de Bovinos de LecheDocumento114 pagineClase de Bovinos de LechemariaNessuna valutazione finora
- 3.1 MielopoyesisDocumento32 pagine3.1 MielopoyesisMichelle MoralesNessuna valutazione finora
- Asma Bronquial Modelo Virginia HendersonDocumento53 pagineAsma Bronquial Modelo Virginia HendersonJose Maria Ramos Cabrera83% (6)
- CirculacionDocumento2 pagineCirculacionRuth VasquezNessuna valutazione finora
- Higiene Postural en El AulaDocumento25 pagineHigiene Postural en El AulaDiana RodriguezNessuna valutazione finora
- Plasma Rico en Plaquetas 1 PDFDocumento17 paginePlasma Rico en Plaquetas 1 PDFGustavoNessuna valutazione finora
- Dolor Pelvico CrónicoDocumento9 pagineDolor Pelvico CrónicoPaulaAlvarezSuaNessuna valutazione finora
- Capitulo 4Documento10 pagineCapitulo 4Luis Ivan MoralesNessuna valutazione finora
- 3.capítulo II. Bases TeóricasDocumento8 pagine3.capítulo II. Bases TeóricasDenissovich JMNessuna valutazione finora
- El Maravilloso Mundo de Los Pokemon PDFDocumento46 pagineEl Maravilloso Mundo de Los Pokemon PDFCarlos Rocha Alvarado50% (2)
- Moraxella BovisDocumento11 pagineMoraxella BovisCristian Vázquez TerrerosNessuna valutazione finora