Potrebbero piacerti anche
- Seeley's Chapter 8 Nervous SystemDocumento100 pagineSeeley's Chapter 8 Nervous SystemChristine Tapawan88% (8)
- Central Nervous System OVL ANA 2103Documento76 pagineCentral Nervous System OVL ANA 2103Baguma MichaelNessuna valutazione finora
- MCB 62 Midterm 1 Study GuideDocumento6 pagineMCB 62 Midterm 1 Study GuideLauren MikeNessuna valutazione finora
- Nervous System Anatomy and Physiology LectureDocumento105 pagineNervous System Anatomy and Physiology LectureButch Dumdum75% (12)
- 4 Physiology Supplement Handout Based On Ganong For TOPNOTCH 21 by Jaffar PinedaDocumento46 pagine4 Physiology Supplement Handout Based On Ganong For TOPNOTCH 21 by Jaffar PinedaAthena Borja100% (3)
- Biological Relevance of Neurotransmitter ReleaseDocumento12 pagineBiological Relevance of Neurotransmitter ReleaseazzaassNessuna valutazione finora
- Badimon Et Al - 2020 - Negative Feedback Control of Neuronal Activity by MicrogliaDocumento41 pagineBadimon Et Al - 2020 - Negative Feedback Control of Neuronal Activity by Microgliakeon.arbabi.altNessuna valutazione finora
- Activity-Dependent Structural and Functional Plasticity of Astrocyte-Neuron InteractionsDocumento20 pagineActivity-Dependent Structural and Functional Plasticity of Astrocyte-Neuron InteractionsAmar JitjNessuna valutazione finora
- Molecular Mechanisms of Neurotransmitter ReleaseDocumento21 pagineMolecular Mechanisms of Neurotransmitter ReleaseNusrat JahanNessuna valutazione finora
- SynapseDocumento45 pagineSynapseroyjason4518Nessuna valutazione finora
- GABA Depolarizes Immature NeuronsDocumento13 pagineGABA Depolarizes Immature NeuronsDaniel ChaguenNessuna valutazione finora
- Stress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityDocumento18 pagineStress Gates An Astrocytic Energy Reservoir To Impair Synaptic PlasticityCony GSNessuna valutazione finora
- Astrocytes in Endocannabinoid Signalling: ReviewDocumento7 pagineAstrocytes in Endocannabinoid Signalling: ReviewHelington CostaNessuna valutazione finora
- Norepinephrine Modulates The Motility of Resting and Activated Microglia Via Different Adrenergic ReceptorsDocumento13 pagineNorepinephrine Modulates The Motility of Resting and Activated Microglia Via Different Adrenergic ReceptorsCarolAlvaradoNessuna valutazione finora
- Aims and Significance: Protti Sesqui R & D - 1Documento5 pagineAims and Significance: Protti Sesqui R & D - 1Jackson ChenNessuna valutazione finora
- AS Bases Celulares DE LAS Oscilaciones Neuronales: UmmaryDocumento8 pagineAS Bases Celulares DE LAS Oscilaciones Neuronales: Ummarydavid alejandro pinzon sanchezNessuna valutazione finora
- Gating + HomeostaticDocumento7 pagineGating + HomeostaticGhea Mae VillasNessuna valutazione finora
- Efficient Generation of Reciprocal Signals by Inhibition: K.khodakhah@einstein - Yu.eduDocumento42 pagineEfficient Generation of Reciprocal Signals by Inhibition: K.khodakhah@einstein - Yu.eduDulce AguirreNessuna valutazione finora
- Ni Hms 437509Documento19 pagineNi Hms 437509diijah678Nessuna valutazione finora
- M HypothalamusDocumento15 pagineM HypothalamusSrilakshmi SridharanNessuna valutazione finora
- G Proteins: Nicotinic Cholinergic ReceptorsDocumento2 pagineG Proteins: Nicotinic Cholinergic ReceptorsKatrina Vea CalaroNessuna valutazione finora
- Neuromuscular JunctionDocumento16 pagineNeuromuscular JunctionSamadshahirNessuna valutazione finora
- NeurotransmittersDocumento7 pagineNeurotransmittersAnn MutisyaNessuna valutazione finora
- Cytokines As Emerging Regulators of Central Nervou - 231113 - 111110Documento12 pagineCytokines As Emerging Regulators of Central Nervou - 231113 - 111110Maurício DallowayNessuna valutazione finora
- Physiology, Neuromuscular Junction of Skeletal MuscleDocumento8 paginePhysiology, Neuromuscular Junction of Skeletal Musclemethmal subasingheNessuna valutazione finora
- Lin 1993Documento13 pagineLin 1993Stress LessNessuna valutazione finora
- Seizure PDFDocumento3 pagineSeizure PDFjustinedaleespedidoNessuna valutazione finora
- TMP 9 DA2Documento11 pagineTMP 9 DA2FrontiersNessuna valutazione finora
- AcetylcholineDocumento50 pagineAcetylcholineEliza GhinitaNessuna valutazione finora
- Dresbach Et AlDocumento17 pagineDresbach Et Alalma ramirezNessuna valutazione finora
- Chapter 1Documento21 pagineChapter 1Chrisantus OkakaNessuna valutazione finora
- Gaba Actions and Ionic Plasticity in Epilepsy: SciencedirectDocumento8 pagineGaba Actions and Ionic Plasticity in Epilepsy: SciencedirectMuhammad Bayu Zohari HutagalungNessuna valutazione finora
- Norepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaDocumento12 pagineNorepinephrine Modulates Single Hypothalamic Arcuate Neurons ViaMariann DuzzNessuna valutazione finora
- 5 GabaDocumento11 pagine5 GabaJose GiovannyNessuna valutazione finora
- Synaptic TransmissionDocumento17 pagineSynaptic TransmissionbnmjgcNessuna valutazione finora
- Long-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumDocumento9 pagineLong-Term Potentiation of Intrinsic Excitability at The Mossy Fiber - Granule Cell Synapse of Rat CerebellumJar JarNessuna valutazione finora
- Introduction To CNS PharmacologyDocumento49 pagineIntroduction To CNS Pharmacologymatchees-gone rogueNessuna valutazione finora
- Computational Models of Neuron-Astrocyte Interaction in EpilepsyDocumento10 pagineComputational Models of Neuron-Astrocyte Interaction in EpilepsyEmma FineNessuna valutazione finora
- Neuro PlasticityDocumento67 pagineNeuro Plasticitydrkadiyala2100% (1)
- As Troc YtesDocumento16 pagineAs Troc YtesMelanie Christine Soriano PascuaNessuna valutazione finora
- Novel Mechanisms of Action of Three Antiepileptic Drugs, Vigabatrin, Tiagabine, and TopiramateDocumento8 pagineNovel Mechanisms of Action of Three Antiepileptic Drugs, Vigabatrin, Tiagabine, and Topiramatedpf050Nessuna valutazione finora
- Velisek 2017 Puritan S View of The Menage A TroisDocumento3 pagineVelisek 2017 Puritan S View of The Menage A TroissilenceindigoNessuna valutazione finora
- Neurotransmission, Measuring Chemical Events In: Advanced ArticleDocumento12 pagineNeurotransmission, Measuring Chemical Events In: Advanced ArticleazzaassNessuna valutazione finora
- AstrocytesDocumento19 pagineAstrocytesMaria PardoNessuna valutazione finora
- BDNF en PCKDocumento8 pagineBDNF en PCKJesusNessuna valutazione finora
- Nociceptive Stimuli Activate The Hypothalamus-HabeDocumento24 pagineNociceptive Stimuli Activate The Hypothalamus-HabeVissente TapiaNessuna valutazione finora
- Cells: The Potential of Targeting Brain Pathology With Ascl1/Mash1Documento7 pagineCells: The Potential of Targeting Brain Pathology With Ascl1/Mash1Paige MunroeNessuna valutazione finora
- Introduction To Neural Plasticity MechanismDocumento9 pagineIntroduction To Neural Plasticity MechanismAlain CésarNessuna valutazione finora
- Central Pattern GeneratorsDocumento4 pagineCentral Pattern GeneratorsCHANGEZ KHAN SARDARNessuna valutazione finora
- HPHY 225 Assignment - FinalDocumento5 pagineHPHY 225 Assignment - FinalSylvester MathiasNessuna valutazione finora
- Varela Parodi 2011Documento4 pagineVarela Parodi 2011Jorge ParodiNessuna valutazione finora
- PIIS2211124723007945Documento23 paginePIIS2211124723007945Murillo MoraesNessuna valutazione finora
- Pathophysiology of EpilepsyDocumento16 paginePathophysiology of EpilepsyYexalen Zazil100% (1)
- Astrocito y SueñoDocumento17 pagineAstrocito y SueñoLEISLY TATIANA OVIEDO GOMEZNessuna valutazione finora
- c30 2014 PannaschDocumento13 paginec30 2014 PannaschkarlosicaNessuna valutazione finora
- KULIAH BLOK SYARAF PERILAKU-1-ELEKTROFISIOLOGI OTAK & EEG - DR SonnyDocumento104 pagineKULIAH BLOK SYARAF PERILAKU-1-ELEKTROFISIOLOGI OTAK & EEG - DR SonnyAndhani putri.kNessuna valutazione finora
- Pocket - Companion - To - Guyton - and - Hall - Textbook - of - Medical - Physiology - ExportedDocumento7 paginePocket - Companion - To - Guyton - and - Hall - Textbook - of - Medical - Physiology - Exportedhassnfiras04Nessuna valutazione finora
- Synaptic Physiology 1Documento16 pagineSynaptic Physiology 1snowyppuppyNessuna valutazione finora
- Applsci 09 04719Documento31 pagineApplsci 09 04719RANUM WANUDYA YUNASNessuna valutazione finora
- Christensen 2016Documento8 pagineChristensen 2016Lina DarkNessuna valutazione finora
- The Circadian System: A Regulatory Feedback Network of Periphery and BrainDocumento12 pagineThe Circadian System: A Regulatory Feedback Network of Periphery and BrainOscarMavilaHerediaNessuna valutazione finora
- Perception Deco PDFDocumento8 paginePerception Deco PDFxmlbioxNessuna valutazione finora
- Margarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisDocumento12 pagineMargarita Zachariou Et Al - Sensory Gating and Its Modulation by Cannabinoids: Electrophysiological, Computational and Mathematical AnalysisJmasnNessuna valutazione finora
- Neurotransmission: Proceedings of the Sixth International Congress of PharmacologyDa EverandNeurotransmission: Proceedings of the Sixth International Congress of PharmacologyLiisa AhteeNessuna valutazione finora
- Neuronal Networks in Brain Function, CNS Disorders, and TherapeuticsDa EverandNeuronal Networks in Brain Function, CNS Disorders, and TherapeuticsCarl FaingoldNessuna valutazione finora
- Fractional Fourier TransformDocumento6 pagineFractional Fourier TransformLucas GallindoNessuna valutazione finora
- Finding Hidden ComponentsDocumento21 pagineFinding Hidden ComponentsLucas GallindoNessuna valutazione finora
- Discrete-Time Fourier TransformDocumento5 pagineDiscrete-Time Fourier TransformLucas GallindoNessuna valutazione finora
- Hadamard TransformDocumento4 pagineHadamard TransformLucas GallindoNessuna valutazione finora
- Disintegration TheoremDocumento3 pagineDisintegration TheoremLucas GallindoNessuna valutazione finora
- Nyquist-Shannon Sampling TheoremDocumento10 pagineNyquist-Shannon Sampling TheoremLucas GallindoNessuna valutazione finora
- Discrete Hartley TransformDocumento5 pagineDiscrete Hartley TransformLucas GallindoNessuna valutazione finora
- Dot ProductDocumento6 pagineDot ProductLucas GallindoNessuna valutazione finora
- Natural Therapies Overview Report FinalDocumento188 pagineNatural Therapies Overview Report FinalLucas GallindoNessuna valutazione finora
- Cavalieri's PrincipleDocumento4 pagineCavalieri's PrincipleLucas GallindoNessuna valutazione finora
- PragPub The Second Iteration - Teaching Kids ProgrammingDocumento30 paginePragPub The Second Iteration - Teaching Kids ProgrammingLucas Gallindo100% (1)
- Symmetry of Second DerivativesDocumento4 pagineSymmetry of Second DerivativesLucas GallindoNessuna valutazione finora
- Fast Fourier TransformDocumento10 pagineFast Fourier TransformLucas GallindoNessuna valutazione finora
- The Costs and Benefits of World-Class UniversitiesDocumento5 pagineThe Costs and Benefits of World-Class UniversitiesLucas GallindoNessuna valutazione finora
- Convolution TheoremDocumento4 pagineConvolution TheoremLucas GallindoNessuna valutazione finora
- Fourier TransformDocumento23 pagineFourier TransformLucas Gallindo100% (1)
- Fubini's TheoremDocumento6 pagineFubini's TheoremLucas GallindoNessuna valutazione finora
- Laplace TransformDocumento13 pagineLaplace TransformLucas GallindoNessuna valutazione finora
- Discrete Fourier TransformDocumento12 pagineDiscrete Fourier TransformLucas GallindoNessuna valutazione finora
- The Statistical Analysis of World Class UniversitiesDocumento21 pagineThe Statistical Analysis of World Class UniversitiesLucas GallindoNessuna valutazione finora
- Creating World-Class Universities in Developing CountriesDocumento16 pagineCreating World-Class Universities in Developing CountriesLucas GallindoNessuna valutazione finora
- Harmonic AnalysisDocumento4 pagineHarmonic AnalysisLucas GallindoNessuna valutazione finora
- Pilates Method For Women's HealthDocumento12 paginePilates Method For Women's HealthLucas GallindoNessuna valutazione finora
- Fun With The Bell CurveDocumento2 pagineFun With The Bell CurveLucas GallindoNessuna valutazione finora
- Information Aggregation, With Application To Monotone Ordering, Advocacy, and ConvivialityDocumento173 pagineInformation Aggregation, With Application To Monotone Ordering, Advocacy, and ConvivialityLucas GallindoNessuna valutazione finora
- Division by ThreeDocumento35 pagineDivision by ThreeLucas GallindoNessuna valutazione finora
- 1207.0580improving Neural Networks by Preventing Co-Adaptation of Feature DetectorsDocumento18 pagine1207.0580improving Neural Networks by Preventing Co-Adaptation of Feature DetectorsLucas GallindoNessuna valutazione finora
- Samsung Galaxy S5 Disassemble GuideDocumento9 pagineSamsung Galaxy S5 Disassemble GuideLucas GallindoNessuna valutazione finora
- Brain Facts Book PDFDocumento96 pagineBrain Facts Book PDFjlangseth100% (1)
- Accepted Manuscript: and BiologyDocumento24 pagineAccepted Manuscript: and BiologylukeNessuna valutazione finora
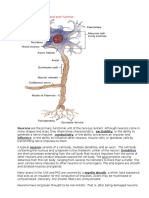
- Nervous Tissue-LabDocumento32 pagineNervous Tissue-LabAlexis PriceNessuna valutazione finora
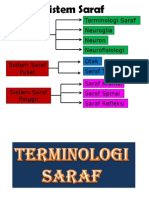
- Tisu Neural Neurofisiologi Neuron Neuroglia Terminologi SarafDocumento141 pagineTisu Neural Neurofisiologi Neuron Neuroglia Terminologi SarafRainne LeeNessuna valutazione finora
- Questions Examens PhysioDocumento32 pagineQuestions Examens PhysioZeineb KammounNessuna valutazione finora
- Histology of Nervous SystemDocumento59 pagineHistology of Nervous SystemMelissa Aina Mohd YusofNessuna valutazione finora
- Atlas NeuroanatomyDocumento874 pagineAtlas NeuroanatomyBilge Yilmaz100% (3)
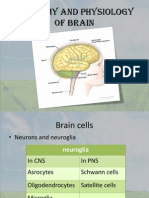
- Anatomy and Physiology of BrainDocumento21 pagineAnatomy and Physiology of BrainNavjot BrarNessuna valutazione finora
- 86 Research Papers Supporting The Vaccine Autism LinkDocumento69 pagine86 Research Papers Supporting The Vaccine Autism LinkDanielPerez100% (1)
- Anatomy & Physiology 1 Chapter 9 The Nervous System Flashcards - QuizletDocumento7 pagineAnatomy & Physiology 1 Chapter 9 The Nervous System Flashcards - Quizletmalenya1100% (1)
- K3. 2014 11 10 Histology of Nervous SystemDocumento84 pagineK3. 2014 11 10 Histology of Nervous SystemRadian Dipta PNessuna valutazione finora
- Chapt12 LectureDocumento197 pagineChapt12 LecturePye Antwan DelvaNessuna valutazione finora
- To Unlock The Brain's Mysteries, Purée It - New York Times Magazine - MediumDocumento10 pagineTo Unlock The Brain's Mysteries, Purée It - New York Times Magazine - MediumSebastian FloresNessuna valutazione finora
- Ch02 PowerPoints FastInternet PDFDocumento33 pagineCh02 PowerPoints FastInternet PDFskdjfNessuna valutazione finora
- Neuro Study GuideDocumento11 pagineNeuro Study GuideEllen Hennings100% (1)
- Glial Reservoir Computing: Liverpool Hope University Liverpool Hope UniversityDocumento6 pagineGlial Reservoir Computing: Liverpool Hope University Liverpool Hope UniversityMonia LandolsiNessuna valutazione finora
- Anatomy and Physiology Chapter 19 Neurological SystemDocumento38 pagineAnatomy and Physiology Chapter 19 Neurological SystemBrantNessuna valutazione finora
- Peripheral Nervous SystemDocumento50 paginePeripheral Nervous SystemaveryNessuna valutazione finora
- Sinelnikov Vol IIIDocumento395 pagineSinelnikov Vol IIIFlorescu Diana NicoletaNessuna valutazione finora
- Progress in Neurodegeneration-The Role of MetalsDocumento358 pagineProgress in Neurodegeneration-The Role of MetalsMaria Rosa100% (2)
- Introduction To NeuroanatomyDocumento49 pagineIntroduction To NeuroanatomyPratik Dhakal100% (1)
- 1 Part I Central Nervous System 4zaDocumento32 pagine1 Part I Central Nervous System 4zaapi-302883249Nessuna valutazione finora
- Histology of Synapse and Neuroglia by Dr. RoomiDocumento18 pagineHistology of Synapse and Neuroglia by Dr. RoomiMudassar Roomi100% (1)
- Cognitive NeuroscienceDocumento42 pagineCognitive NeuroscienceDiya Saher100% (1)
- 30 Scientific Studies That Demonstrate Vaccines Can Cause AutismDocumento23 pagine30 Scientific Studies That Demonstrate Vaccines Can Cause Autismazrulelias100% (6)