Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- AI Intelligence (Sam Charrington and Abeba Birhane)Documento2 pagineAI Intelligence (Sam Charrington and Abeba Birhane)Ethel Shammah Waoulda Acleta90% (10)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- AR 700-84 (Issue and Sale of Personal Clothing)Documento96 pagineAR 700-84 (Issue and Sale of Personal Clothing)ncfranklinNessuna valutazione finora
- Listo Si Kap!Documento32 pagineListo Si Kap!Bluboy100% (3)
- Names of PartnerDocumento7 pagineNames of PartnerDana-Zaza BajicNessuna valutazione finora
- Book Item 97952Documento18 pagineBook Item 97952Shairah May MendezNessuna valutazione finora
- Judgments of Adminstrative LawDocumento22 pagineJudgments of Adminstrative Lawpunit gaurNessuna valutazione finora
- The Absent Presence of Progressive Rock in The British Music Press 1968 1974 PDFDocumento33 pagineThe Absent Presence of Progressive Rock in The British Music Press 1968 1974 PDFwago_itNessuna valutazione finora
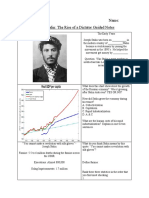
- Stalin Guided NotesDocumento2 pagineStalin Guided Notesapi-536892868Nessuna valutazione finora
- Assets Misappropriation in The Malaysian Public AnDocumento5 pagineAssets Misappropriation in The Malaysian Public AnRamadona SimbolonNessuna valutazione finora
- Mock Act 1 - Student - 2023Documento8 pagineMock Act 1 - Student - 2023Big bundahNessuna valutazione finora
- Travel Reservation August 18 For FREDI ISWANTODocumento2 pagineTravel Reservation August 18 For FREDI ISWANTOKasmi MinukNessuna valutazione finora
- AssignmentDocumento25 pagineAssignmentPrashan Shaalin FernandoNessuna valutazione finora
- Bond by A Person Obtaining Letters of Administration With Two SuretiesDocumento2 pagineBond by A Person Obtaining Letters of Administration With Two Suretiessamanta pandeyNessuna valutazione finora
- BRUNOISEDocumento1 paginaBRUNOISEMelchor Dagandan Toylo JfyNessuna valutazione finora
- Alpha Beta Gamma Delta Epsilon Zeta Eta Theta Iota Kappa Lambda MuDocumento3 pagineAlpha Beta Gamma Delta Epsilon Zeta Eta Theta Iota Kappa Lambda MulanNessuna valutazione finora
- The Mental Health Act 2018 Tested by High Court of UgandaDocumento7 pagineThe Mental Health Act 2018 Tested by High Court of UgandaLDC Online ResourcesNessuna valutazione finora
- Đề Cương CK - QuestionsDocumento2 pagineĐề Cương CK - QuestionsDiệu Phương LêNessuna valutazione finora
- About ArevaDocumento86 pagineAbout ArevaAbhinav TyagiNessuna valutazione finora
- Appleby, Telling The Truth About History, Introduction and Chapter 4Documento24 pagineAppleby, Telling The Truth About History, Introduction and Chapter 4Steven LubarNessuna valutazione finora
- Tucker Travis-Reel Site 2013-1Documento2 pagineTucker Travis-Reel Site 2013-1api-243050578Nessuna valutazione finora
- Code of Civil Procedure: Recording of Evidence by Court/Court CommissionerDocumento12 pagineCode of Civil Procedure: Recording of Evidence by Court/Court CommissionercahirenrajaNessuna valutazione finora
- Ingles Junio 17 CanariasDocumento5 pagineIngles Junio 17 CanariaskinkirevolutionNessuna valutazione finora
- Ais CH5Documento30 pagineAis CH5MosabAbuKhater100% (1)
- People Vs SolangonDocumento7 paginePeople Vs SolangonChris YumNessuna valutazione finora
- Anderson v. Eighth Judicial District Court - OpinionDocumento8 pagineAnderson v. Eighth Judicial District Court - OpinioniX i0Nessuna valutazione finora
- A Documentary Report of Work Immersion Undertaken at The City Mayor'S OfficeDocumento17 pagineA Documentary Report of Work Immersion Undertaken at The City Mayor'S OfficeHamdan AbisonNessuna valutazione finora
- Tiktok PresentationDocumento17 pagineTiktok Presentationapi-681531475Nessuna valutazione finora
- Personal Data Breach Notifications in The GDPRDocumento17 paginePersonal Data Breach Notifications in The GDPRgschandel3356Nessuna valutazione finora
- Balkan Languages - Victor FriedmanDocumento12 pagineBalkan Languages - Victor FriedmanBasiol Chulev100% (1)
- Company Analysis of Axis BankDocumento66 pagineCompany Analysis of Axis BankKirti Bhite100% (1)