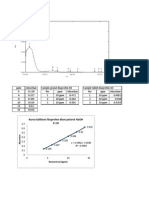
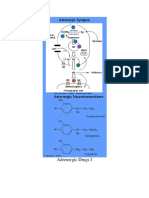
Potrebbero piacerti anche
- Success Factors for Fish Larval ProductionDa EverandSuccess Factors for Fish Larval ProductionLuis ConceicaoNessuna valutazione finora
- Quercetin: Toxicology CarcinogenesisDocumento171 pagineQuercetin: Toxicology CarcinogenesisPig NguyenNessuna valutazione finora
- The Cleveland Clinic Manual of Dynamic Endocrine TestingDa EverandThe Cleveland Clinic Manual of Dynamic Endocrine TestingNessuna valutazione finora
- Tacrine Induce Hepatotoksik PDFDocumento9 pagineTacrine Induce Hepatotoksik PDFItamahYulaikhaNessuna valutazione finora
- LifeWave X39 Pilot Demuestra Cambios Ligeros ActivadosDocumento12 pagineLifeWave X39 Pilot Demuestra Cambios Ligeros Activadosave_fenix_mileniumNessuna valutazione finora
- Thiabendazole (WHO Food Additives Series 39)Documento7 pagineThiabendazole (WHO Food Additives Series 39)hellen86150Nessuna valutazione finora
- Fetal Growth and Reproductive Performance in Ewes Administered GNRH Agonist On Day 12 Post-MatingDocumento10 pagineFetal Growth and Reproductive Performance in Ewes Administered GNRH Agonist On Day 12 Post-MatingrevinaNessuna valutazione finora
- Pengaruh Makanan Pada CaptoprilDocumento5 paginePengaruh Makanan Pada CaptoprilNiluh putu Satria maharaniNessuna valutazione finora
- Artículo BiodisponibilidadDocumento9 pagineArtículo BiodisponibilidadLuis Enrique SanchezNessuna valutazione finora
- Rosenkrantz 1988Documento12 pagineRosenkrantz 1988NoemiNessuna valutazione finora
- Anthropometrical Parameters and Markers of Obesity in RatsDocumento9 pagineAnthropometrical Parameters and Markers of Obesity in RatsSammer BurgosNessuna valutazione finora
- Latham 1999Documento6 pagineLatham 1999Juci StoregjerdeNessuna valutazione finora
- Effect of Ovariectomy On The Levels of Plasma Sex Hormones in Albino Rats E. A. Alagwu., R. O. NneliDocumento5 pagineEffect of Ovariectomy On The Levels of Plasma Sex Hormones in Albino Rats E. A. Alagwu., R. O. NneliCaroline AgathaNessuna valutazione finora
- Melamine: 1.1 Chemical and Physical DataDocumento10 pagineMelamine: 1.1 Chemical and Physical DataKatrien VandenheckeNessuna valutazione finora
- 84 - Ijbpr 2011, 2 (2), 80-84Documento5 pagine84 - Ijbpr 2011, 2 (2), 80-84Sujith KuttanNessuna valutazione finora
- Butyl Benzil PhthalateDocumento15 pagineButyl Benzil PhthalateLALUKISNessuna valutazione finora
- Chlorodifluoromethane CASRN 75-45-6: IRIS Assessment Development ProcessDocumento12 pagineChlorodifluoromethane CASRN 75-45-6: IRIS Assessment Development ProcessridwanNessuna valutazione finora
- Paracetamol: 1.1 Chemical and Physical DataDocumento49 pagineParacetamol: 1.1 Chemical and Physical DataKimskiusNessuna valutazione finora
- Bioequivalence Study of Carbamazepine Tablets in Vitro in Vivo CorrelationDocumento8 pagineBioequivalence Study of Carbamazepine Tablets in Vitro in Vivo CorrelationFriska Aprianti100% (1)
- The Pathological Features of CyclophosphDocumento5 pagineThe Pathological Features of Cyclophosphissamlfc92Nessuna valutazione finora
- In Vitro Evaluation of The Pollen Extract Cernitin T 60Documento5 pagineIn Vitro Evaluation of The Pollen Extract Cernitin T 60api-442513573Nessuna valutazione finora
- Pharmacokinetics and Dose Selection of A Novel, Long-Acting Transdermal Fentanyl Solution in Healthy Laboratory BeaglesDocumento6 paginePharmacokinetics and Dose Selection of A Novel, Long-Acting Transdermal Fentanyl Solution in Healthy Laboratory BeaglesRhanna AbreuNessuna valutazione finora
- The Open Natural Products Journal, 2009, 2, 68-70Documento3 pagineThe Open Natural Products Journal, 2009, 2, 68-70Valentina AnutaNessuna valutazione finora
- MALONDIALDEHYDE - IARC (1985) and The Compound Was Classified in IARCDocumento11 pagineMALONDIALDEHYDE - IARC (1985) and The Compound Was Classified in IARCPritta TaradipaNessuna valutazione finora
- AZACITIDINEDocumento17 pagineAZACITIDINEjian_wyuNessuna valutazione finora
- Melatonine 2Documento5 pagineMelatonine 2unrealragdollsNessuna valutazione finora
- Cachexia-Inducing (MAC16) : Effect of Megestrol Weight by Factor and MiceDocumento5 pagineCachexia-Inducing (MAC16) : Effect of Megestrol Weight by Factor and MicedonkeyendutNessuna valutazione finora
- Thibault 2019Documento8 pagineThibault 2019Mafe HerreraNessuna valutazione finora
- Toxicological, Pathological, Teratological Studies in Animals With CephradineDocumento4 pagineToxicological, Pathological, Teratological Studies in Animals With CephradinePharmacistNessuna valutazione finora
- Brjcancer00059 0200Documento6 pagineBrjcancer00059 0200irene hoffmannNessuna valutazione finora
- Ccras and NinDocumento9 pagineCcras and NinPRAKASH DESHPANDENessuna valutazione finora
- Ssri On CancerDocumento7 pagineSsri On CancerArpit SanghaniNessuna valutazione finora
- A Comparative Study of The Gastrointestinal of A Pellet and Tablet Formulation TransitDocumento11 pagineA Comparative Study of The Gastrointestinal of A Pellet and Tablet Formulation TransitFarmasi Kelas ANessuna valutazione finora
- Aac00042 0048Documento5 pagineAac00042 0048Mohamed Abo El FadlNessuna valutazione finora
- 3057 FullDocumento7 pagine3057 FullAnonymous Wcj4C3jNessuna valutazione finora
- Adverse Effects of Incorporating Ketoprofen Into Established Rodent StudiesDocumento11 pagineAdverse Effects of Incorporating Ketoprofen Into Established Rodent StudiesVivi FatimatuzzuhroNessuna valutazione finora
- A 90-Day Subchronic Toxicity Study of Neem Oil, A Azadirachta Indica Oil, in MiceDocumento11 pagineA 90-Day Subchronic Toxicity Study of Neem Oil, A Azadirachta Indica Oil, in Micehenry72Nessuna valutazione finora
- Comparison of The of Netilmicin and Gentamicin: NephrotoxicityDocumento5 pagineComparison of The of Netilmicin and Gentamicin: NephrotoxicityCLPHtheoryNessuna valutazione finora
- ContentServer - Asp 13Documento7 pagineContentServer - Asp 13Fitria NurulfathNessuna valutazione finora
- เห็ดการบูรกับมะเร็งตับ8Documento8 pagineเห็ดการบูรกับมะเร็งตับ8patarapen.mNessuna valutazione finora
- Diabetes PDFDocumento12 pagineDiabetes PDFDANIELA MALAGÓN MONTAÑONessuna valutazione finora
- Pharmacokinetics of Ceftriaxone in Buffalo Calves (Bubalus Bubalis) Following Intravenous and Intramuscular AdministrationDocumento5 paginePharmacokinetics of Ceftriaxone in Buffalo Calves (Bubalus Bubalis) Following Intravenous and Intramuscular AdministrationBao TranNessuna valutazione finora
- Guidelines For Toxicity/safety Profile Evaluation of Ayurved & Siddha, Plant DrugsDocumento5 pagineGuidelines For Toxicity/safety Profile Evaluation of Ayurved & Siddha, Plant DrugsPRAKASH DESHPANDENessuna valutazione finora
- 2012 82 6 3Documento12 pagine2012 82 6 3Dyego CarléttiNessuna valutazione finora
- Me Hanna 2018Documento36 pagineMe Hanna 2018Mtra. Mayra N. Elizalde VillarrealNessuna valutazione finora
- Studies On The Pharmacokinetics and Pharmacodynamics of Mirtazapine in Healthy Young CatsDocumento9 pagineStudies On The Pharmacokinetics and Pharmacodynamics of Mirtazapine in Healthy Young CatsWilliam ChandlerNessuna valutazione finora
- Efficacy and Safety of Monascus Purpureus Went Rice in Subjects With HyperlipidemiaDocumento8 pagineEfficacy and Safety of Monascus Purpureus Went Rice in Subjects With HyperlipidemiaAyshe SlocumNessuna valutazione finora
- Ijms 19 03457Documento6 pagineIjms 19 03457efendirahimNessuna valutazione finora
- Effect of Soybeans On Lipid Profile of Female and Male Albino RatsDocumento9 pagineEffect of Soybeans On Lipid Profile of Female and Male Albino RatsAdib MustofaNessuna valutazione finora
- Effects of Dystocia, Fetotomy and Caesarian Sectios On The Liver EnzymesDocumento8 pagineEffects of Dystocia, Fetotomy and Caesarian Sectios On The Liver EnzymesiggyNessuna valutazione finora
- Aftereffect of Gemcitabine Around The Amount of Sideline Blood Lymphocyte Autophagy in Kids Along With Endemic Lupus ErythematosusqsulpDocumento2 pagineAftereffect of Gemcitabine Around The Amount of Sideline Blood Lymphocyte Autophagy in Kids Along With Endemic Lupus Erythematosusqsulpcelerychive5Nessuna valutazione finora
- Environmental Toxicology - 2013 - Lan - Triclosan Exhibits A Tendency To Accumulate in The Epididymis and Shows Sperm-1Documento9 pagineEnvironmental Toxicology - 2013 - Lan - Triclosan Exhibits A Tendency To Accumulate in The Epididymis and Shows Sperm-1Sultansarwar SarwarNessuna valutazione finora
- Hyperlipidemia Induced by Triton WR1339 (Tyloxapol) in Wistar RatsDocumento3 pagineHyperlipidemia Induced by Triton WR1339 (Tyloxapol) in Wistar RatsdonNessuna valutazione finora
- 3442 FullDocumento7 pagine3442 FullAbigail PheiliaNessuna valutazione finora
- The Effects of Sub-Chronic Exposure of Wistar Rats To The Herbicide Glyphosate-BiocarbDocumento6 pagineThe Effects of Sub-Chronic Exposure of Wistar Rats To The Herbicide Glyphosate-BiocarbFreddy A. ManayayNessuna valutazione finora
- CimetidineDocumento3 pagineCimetidineRodolfo Lone UlvenNessuna valutazione finora
- PROPYLTHIOURACILDocumento35 paginePROPYLTHIOURACILMagdy Ali ELsherbenyNessuna valutazione finora
- Vivra Tucson Study SheetsDocumento58 pagineVivra Tucson Study SheetsRobert Davidson, M.D., Ph.D.Nessuna valutazione finora
- Giammanco M. Effects of 3,5-Diiodo-L-Thyronine On The Liver of High Fat Diet Fed Rats. JBR 2016Documento6 pagineGiammanco M. Effects of 3,5-Diiodo-L-Thyronine On The Liver of High Fat Diet Fed Rats. JBR 2016Marco GiammancoNessuna valutazione finora
- Kukanich 2020Documento10 pagineKukanich 2020Gianne K. SantosNessuna valutazione finora
- Glukosa DialabDocumento2 pagineGlukosa DialabDian Ayu UtamiNessuna valutazione finora
- A3 Nomor (Vinil Doff)Documento1 paginaA3 Nomor (Vinil Doff)Dian Ayu UtamiNessuna valutazione finora
- Serial Number Key Avg CRACKDocumento1 paginaSerial Number Key Avg CRACKDian Ayu UtamiNessuna valutazione finora
- NOMOR ABU2-dikonversiDocumento1 paginaNOMOR ABU2-dikonversiDian Ayu UtamiNessuna valutazione finora
- Urea DialabDocumento2 pagineUrea DialabDian Ayu UtamiNessuna valutazione finora
- WWW - Diameb.ua Manuals Eng D95595Documento2 pagineWWW - Diameb.ua Manuals Eng D95595Dian Ayu UtamiNessuna valutazione finora
- Zovirax Cold Sore CreamDocumento3 pagineZovirax Cold Sore CreamDian Ayu UtamiNessuna valutazione finora
- Anti-Inflammatory, Antioxidant and Anticancer Activity of Quercetin and Its AnaloguesDocumento11 pagineAnti-Inflammatory, Antioxidant and Anticancer Activity of Quercetin and Its AnaloguesDian Ayu UtamiNessuna valutazione finora
- Kurva Kalibrasi Ibuprofen Dlam Pelarut Naoh 0.1N: Konsentrasi (PPM)Documento3 pagineKurva Kalibrasi Ibuprofen Dlam Pelarut Naoh 0.1N: Konsentrasi (PPM)Dian Ayu UtamiNessuna valutazione finora
- Jen-Hung Yang, Te-Chun Hsia, Hsiu-Maan Kuo, Pei-Dawn Lee Chao, Chi-Chung Chou, Yau-Huei Wei, and Jing-Gung ChungDocumento9 pagineJen-Hung Yang, Te-Chun Hsia, Hsiu-Maan Kuo, Pei-Dawn Lee Chao, Chi-Chung Chou, Yau-Huei Wei, and Jing-Gung ChungDian Ayu UtamiNessuna valutazione finora
- Iodin PDFDocumento3 pagineIodin PDFDian Ayu UtamiNessuna valutazione finora
- Microbiol. Mol. Biol. Rev. 1998 Rao 597 635Documento40 pagineMicrobiol. Mol. Biol. Rev. 1998 Rao 597 635Dian Ayu UtamiNessuna valutazione finora
- QualitatiQualitative and Quantitative of Herbal Medicine STD - Pdfve and Quantitative of Herbal Medicine STDDocumento30 pagineQualitatiQualitative and Quantitative of Herbal Medicine STD - Pdfve and Quantitative of Herbal Medicine STDDian Ayu UtamiNessuna valutazione finora
- Application of Chitosan To Modulate Hpmc-Ec Based ControlledDocumento10 pagineApplication of Chitosan To Modulate Hpmc-Ec Based ControlledDian Ayu UtamiNessuna valutazione finora
- Parasites E.histolitycaDocumento3 pagineParasites E.histolitycaDian Ayu UtamiNessuna valutazione finora
- Form Pendaftaran DhisDocumento11 pagineForm Pendaftaran DhisDian Ayu UtamiNessuna valutazione finora
- Lampiran Kfa 2 KCKTDocumento2 pagineLampiran Kfa 2 KCKTDian Ayu UtamiNessuna valutazione finora
- Parasites CDC TripanosomiasisDocumento5 pagineParasites CDC TripanosomiasisDian Ayu UtamiNessuna valutazione finora
- Astra 1st Development Program 2013Documento1 paginaAstra 1st Development Program 2013Dian Ayu UtamiNessuna valutazione finora
- Adrenergic Drugs IDocumento11 pagineAdrenergic Drugs IDian Ayu UtamiNessuna valutazione finora
- The Seven Crystal SystemsDocumento2 pagineThe Seven Crystal SystemsDian Ayu UtamiNessuna valutazione finora
- Basic InformationDocumento18 pagineBasic InformationDian Ayu UtamiNessuna valutazione finora
- Ajac20100100003 89387705 PDFDocumento3 pagineAjac20100100003 89387705 PDFDian Ayu UtamiNessuna valutazione finora
- Chuyên Đề ConjunctionDocumento5 pagineChuyên Đề ConjunctionKhánh Linh TrịnhNessuna valutazione finora
- Caption Sheet 4-Kailynn BDocumento4 pagineCaption Sheet 4-Kailynn Bapi-549116310Nessuna valutazione finora
- Bluforest, Inc. (OTC: BLUF) InvestigationDocumento5 pagineBluforest, Inc. (OTC: BLUF) Investigationfraudinstitute100% (1)
- 1 Quarter: 5 Grade Benchmark Scope and Sequence Pacing Guide 2015-2016Documento3 pagine1 Quarter: 5 Grade Benchmark Scope and Sequence Pacing Guide 2015-2016api-292307509Nessuna valutazione finora
- Chemistry NotesDocumento11 pagineChemistry Notesraifaisal9267% (12)
- 5 Years High and Low PointsDocumento7 pagine5 Years High and Low PointsNaresh Kumar VishwakarmaNessuna valutazione finora
- EET - Formulas - Christmas TermDocumento3 pagineEET - Formulas - Christmas TermJMDNessuna valutazione finora
- Double-Outlet Right Ventricle With An An Intact Interventricular Septum and Concurrent Hypoplastic Left Ventricle in A CalfDocumento6 pagineDouble-Outlet Right Ventricle With An An Intact Interventricular Septum and Concurrent Hypoplastic Left Ventricle in A CalfYoga RivaldiNessuna valutazione finora
- FREEWAT Vol0 v.1.1.2Documento159 pagineFREEWAT Vol0 v.1.1.2Jonathan QuirozNessuna valutazione finora
- Logistics Operation PlanningDocumento25 pagineLogistics Operation PlanningLeonard AntoniusNessuna valutazione finora
- Tech Manual 1396 Rev. B: 3.06/4.06" 15,000 Psi ES BOPDocumento39 pagineTech Manual 1396 Rev. B: 3.06/4.06" 15,000 Psi ES BOPEl Mundo De Yosed100% (1)
- 1.1 - Selectividad Acti9Documento34 pagine1.1 - Selectividad Acti9Bladimir MichelNessuna valutazione finora
- Manual de Taller sk350 PDFDocumento31 pagineManual de Taller sk350 PDFLeo Perez100% (1)
- 2606 PDFDocumento6 pagine2606 PDFzainab jehangirNessuna valutazione finora
- Macleod - 1974 - Lucian's Knowledge of TheophrastusDocumento2 pagineMacleod - 1974 - Lucian's Knowledge of TheophrastusSIMONE BLAIRNessuna valutazione finora
- Julie RiesenweberDocumento14 pagineJulie RiesenweberisidorabilbaoNessuna valutazione finora
- Fluid Solids Operations: High HighDocumento20 pagineFluid Solids Operations: High HighPriscilaPrzNessuna valutazione finora
- WRP Examination Extension 1 2012Documento12 pagineWRP Examination Extension 1 2012Belinda JadeNessuna valutazione finora
- Novel Image Enhancement Technique Using CLAHE and Wavelet TransformsDocumento5 pagineNovel Image Enhancement Technique Using CLAHE and Wavelet TransformsInnovative Research PublicationsNessuna valutazione finora
- Welding Research Council: BulletinDocumento28 pagineWelding Research Council: BulletinRogerio Tropia GranjaNessuna valutazione finora
- Web+Presentation+12+July+2016 EA+-+Eric+LumeDocumento57 pagineWeb+Presentation+12+July+2016 EA+-+Eric+LumetranthabinNessuna valutazione finora
- Magneto-Convective Non-Newtonian Nanofluid With Momentum and Temperature Dependent Slip Flow From A Permeable Stretching Sheet With Porous Medium and Chemical ReactionDocumento18 pagineMagneto-Convective Non-Newtonian Nanofluid With Momentum and Temperature Dependent Slip Flow From A Permeable Stretching Sheet With Porous Medium and Chemical ReactionIOSRjournalNessuna valutazione finora
- Resource Access ControlDocumento19 pagineResource Access Controlusamadar707Nessuna valutazione finora
- InTech-Batteries Charging Systems For Electric and Plug in Hybrid Electric VehiclesDocumento20 pagineInTech-Batteries Charging Systems For Electric and Plug in Hybrid Electric VehiclesM VetriselviNessuna valutazione finora
- Electric Bike Monitoring and Controlling System Based On Internet of ThingsDocumento8 pagineElectric Bike Monitoring and Controlling System Based On Internet of ThingsEarl PhamtonhiveNessuna valutazione finora
- Article Unleashing The Power of Your StoryDocumento17 pagineArticle Unleashing The Power of Your StoryAnkit ChhabraNessuna valutazione finora
- 2003 Nissan Altima 2.5 Serivce Manual RSUDocumento12 pagine2003 Nissan Altima 2.5 Serivce Manual RSUAndy Dellinger100% (1)
- Module 12. Big Issues Lesson 12a. Reading. Pages 140-141: No Words TranslationDocumento4 pagineModule 12. Big Issues Lesson 12a. Reading. Pages 140-141: No Words TranslationLeonardo Perez AlegriaNessuna valutazione finora
- Good Stuff Courage, Resilience, Gratitude, Generosity, Forgiveness, and Sacrifice (Akhtar, Salman)Documento221 pagineGood Stuff Courage, Resilience, Gratitude, Generosity, Forgiveness, and Sacrifice (Akhtar, Salman)madhugaggar999Nessuna valutazione finora
- Modular Air Handling Unit (FMA)Documento3 pagineModular Air Handling Unit (FMA)Salamet TeguhNessuna valutazione finora
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouDa EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouValutazione: 4.5 su 5 stelle4.5/5 (62)
- Why We Die: The New Science of Aging and the Quest for ImmortalityDa EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityValutazione: 4 su 5 stelle4/5 (3)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisDa EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisValutazione: 3.5 su 5 stelle3.5/5 (2)
- Tales from Both Sides of the Brain: A Life in NeuroscienceDa EverandTales from Both Sides of the Brain: A Life in NeuroscienceValutazione: 3 su 5 stelle3/5 (18)
- Gut: the new and revised Sunday Times bestsellerDa EverandGut: the new and revised Sunday Times bestsellerValutazione: 4 su 5 stelle4/5 (392)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedDa EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedValutazione: 4 su 5 stelle4/5 (11)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceDa EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceValutazione: 4.5 su 5 stelle4.5/5 (516)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Da EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Valutazione: 4 su 5 stelle4/5 (378)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessDa Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessValutazione: 4 su 5 stelle4/5 (33)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionDa EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionValutazione: 4 su 5 stelle4/5 (811)
- Inside of a Dog: What Dogs See, Smell, and KnowDa EverandInside of a Dog: What Dogs See, Smell, and KnowValutazione: 4 su 5 stelle4/5 (390)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsDa EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsValutazione: 4 su 5 stelle4/5 (5)
- Masterminds: Genius, DNA, and the Quest to Rewrite LifeDa EverandMasterminds: Genius, DNA, and the Quest to Rewrite LifeNessuna valutazione finora
- Human: The Science Behind What Makes Your Brain UniqueDa EverandHuman: The Science Behind What Makes Your Brain UniqueValutazione: 3.5 su 5 stelle3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperDa EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperValutazione: 4.5 su 5 stelle4.5/5 (15)
- Good Without God: What a Billion Nonreligious People Do BelieveDa EverandGood Without God: What a Billion Nonreligious People Do BelieveValutazione: 4 su 5 stelle4/5 (66)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorDa EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNessuna valutazione finora
- Crypt: Life, Death and Disease in the Middle Ages and BeyondDa EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondValutazione: 4 su 5 stelle4/5 (4)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomDa EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomValutazione: 4 su 5 stelle4/5 (215)
- Who's in Charge?: Free Will and the Science of the BrainDa EverandWho's in Charge?: Free Will and the Science of the BrainValutazione: 4 su 5 stelle4/5 (65)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesDa EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesValutazione: 4.5 su 5 stelle4.5/5 (397)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldDa EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldValutazione: 4.5 su 5 stelle4.5/5 (18)
- The Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineDa EverandThe Second Brain: A Groundbreaking New Understanding of Nervous Disorders of the Stomach and IntestineValutazione: 4 su 5 stelle4/5 (17)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemDa EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemValutazione: 4.5 su 5 stelle4.5/5 (115)
- The Invention of Tomorrow: A Natural History of ForesightDa EverandThe Invention of Tomorrow: A Natural History of ForesightValutazione: 4.5 su 5 stelle4.5/5 (5)