Potrebbero piacerti anche
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeDa EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeValutazione: 4 su 5 stelle4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreDa EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreValutazione: 4 su 5 stelle4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItDa EverandNever Split the Difference: Negotiating As If Your Life Depended On ItValutazione: 4.5 su 5 stelle4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceDa EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceValutazione: 4 su 5 stelle4/5 (895)
- Grit: The Power of Passion and PerseveranceDa EverandGrit: The Power of Passion and PerseveranceValutazione: 4 su 5 stelle4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeDa EverandShoe Dog: A Memoir by the Creator of NikeValutazione: 4.5 su 5 stelle4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersDa EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersValutazione: 4.5 su 5 stelle4.5/5 (344)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureDa EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureValutazione: 4.5 su 5 stelle4.5/5 (474)
- Her Body and Other Parties: StoriesDa EverandHer Body and Other Parties: StoriesValutazione: 4 su 5 stelle4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)Da EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Valutazione: 4.5 su 5 stelle4.5/5 (120)
- The Emperor of All Maladies: A Biography of CancerDa EverandThe Emperor of All Maladies: A Biography of CancerValutazione: 4.5 su 5 stelle4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingDa EverandThe Little Book of Hygge: Danish Secrets to Happy LivingValutazione: 3.5 su 5 stelle3.5/5 (399)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyDa EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyValutazione: 3.5 su 5 stelle3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)Da EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Valutazione: 4 su 5 stelle4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaDa EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaValutazione: 4.5 su 5 stelle4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryDa EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryValutazione: 3.5 su 5 stelle3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnDa EverandTeam of Rivals: The Political Genius of Abraham LincolnValutazione: 4.5 su 5 stelle4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealDa EverandOn Fire: The (Burning) Case for a Green New DealValutazione: 4 su 5 stelle4/5 (73)
- The Unwinding: An Inner History of the New AmericaDa EverandThe Unwinding: An Inner History of the New AmericaValutazione: 4 su 5 stelle4/5 (45)
- Sewage Treatmant Plant ManualDocumento51 pagineSewage Treatmant Plant ManualAnonymous uTO1WI100% (2)
- APURE英文2 0Documento14 pagineAPURE英文2 0CHRISTIAN FIDEL VILCA OJEDA0% (1)
- Pwps Mobilepac BrochureDocumento6 paginePwps Mobilepac BrochureMarcos Huata100% (1)
- Criticial Utilities Qualifcation Part IIDocumento12 pagineCriticial Utilities Qualifcation Part IInavas1972Nessuna valutazione finora
- Denah LokerDocumento6 pagineDenah LokerSunflowerNessuna valutazione finora
- Pertanyaan Training Iso 9001Documento1 paginaPertanyaan Training Iso 9001SunflowerNessuna valutazione finora
- Presentation Title: Subheading Goes HereDocumento2 paginePresentation Title: Subheading Goes HereSunflowerNessuna valutazione finora
- Kitinase Dari JamurDocumento7 pagineKitinase Dari JamurSunflowerNessuna valutazione finora
- Alfabet InternasionalDocumento1 paginaAlfabet InternasionalSunflowerNessuna valutazione finora
- Jurnal BrezinskaDocumento8 pagineJurnal BrezinskaSunflowerNessuna valutazione finora
- EnrichmentDocumento9 pagineEnrichmentSunflowerNessuna valutazione finora
- From Lonar LakeDocumento7 pagineFrom Lonar LakeSunflowerNessuna valutazione finora
- Isolasi Dan Seleksi Bakteri Kitinolitik Isolat Lokal Yang Berpotensi Untuk Mengendalikan Larva Nyamuk Aedes Aegypti LDocumento4 pagineIsolasi Dan Seleksi Bakteri Kitinolitik Isolat Lokal Yang Berpotensi Untuk Mengendalikan Larva Nyamuk Aedes Aegypti LSunflowerNessuna valutazione finora
- LowryDocumento15 pagineLowrySunflowerNessuna valutazione finora
- Cassava Characteristics - Ibwebsite (English)Documento9 pagineCassava Characteristics - Ibwebsite (English)SunflowerNessuna valutazione finora
- Jurnal InternasionalDocumento7 pagineJurnal InternasionalSunflowerNessuna valutazione finora
- CinderellaDocumento1 paginaCinderellaRiNessuna valutazione finora
- Supervised Classification of Scatterers On Sar Imaging Based On Incoherent Polarimetric Time-Frequency SignaturesDocumento5 pagineSupervised Classification of Scatterers On Sar Imaging Based On Incoherent Polarimetric Time-Frequency SignaturesSunflowerNessuna valutazione finora
- Bay of BengalDocumento4 pagineBay of Bengalnk4j3tk34j5Nessuna valutazione finora
- List Capel SPK2 1075Documento5 pagineList Capel SPK2 1075singki bernando junitra singki bernando junitraNessuna valutazione finora
- TB Evolution TDAR-TUARDocumento2 pagineTB Evolution TDAR-TUARPrieto RamiroNessuna valutazione finora
- Oman - Strucure and GulfDocumento34 pagineOman - Strucure and GulfAhmed AL Duwaiki100% (2)
- Washer-Disinfectors For Hospitals and Central Sterilisation DepartmentsDocumento72 pagineWasher-Disinfectors For Hospitals and Central Sterilisation DepartmentsRizky ZulfadlanNessuna valutazione finora
- Hydraulic GradientDocumento1 paginaHydraulic GradientPika ThoidingjamNessuna valutazione finora
- Oil Spill Research PaperDocumento5 pagineOil Spill Research Paperc9snjtdx100% (1)
- Brgy-DTP-Forms Hilongos - CBMSDocumento26 pagineBrgy-DTP-Forms Hilongos - CBMSJaypee BucatcatNessuna valutazione finora
- Constant Chlor MC4-50 and MC4-150 Operation and Installation ManualDocumento76 pagineConstant Chlor MC4-50 and MC4-150 Operation and Installation ManualChristian NdembiNessuna valutazione finora
- ANSWER To QUESTION 1 Seven Up Bottling Company Currently Produces The Following ProductsDocumento4 pagineANSWER To QUESTION 1 Seven Up Bottling Company Currently Produces The Following ProductsAbanum GodspowerNessuna valutazione finora
- 2 Company Profile SPP Pumps - Oil & Gas - BrochureDocumento10 pagine2 Company Profile SPP Pumps - Oil & Gas - Brochurelemuel bacsaNessuna valutazione finora
- Pengaruh Implementasi Sistem Irigasi Big Gun Sprinkler Dan Bahan Organik Terhadap Kelengasan Tanah Dan Produksi Jagung Di Lahan KeringDocumento10 paginePengaruh Implementasi Sistem Irigasi Big Gun Sprinkler Dan Bahan Organik Terhadap Kelengasan Tanah Dan Produksi Jagung Di Lahan KeringDonny Nugroho KalbuadiNessuna valutazione finora
- Waterchar IwasteDocumento8 pagineWaterchar IwasteCharisma CabanogNessuna valutazione finora
- Tunnel PresentationDocumento68 pagineTunnel PresentationCatalin Boroianu100% (2)
- Philippine National Standard: Agricultural Structures - Lairage - RuminantsDocumento24 paginePhilippine National Standard: Agricultural Structures - Lairage - RuminantsAlfredo CondeNessuna valutazione finora
- Partition Coefficient EditedDocumento17 paginePartition Coefficient EditedcoltalbNessuna valutazione finora
- Mobilgear 630 MSDS 영문Documento6 pagineMobilgear 630 MSDS 영문thisbaboNessuna valutazione finora
- UBSRSI Environmental ImpactDocumento52 pagineUBSRSI Environmental ImpactJon CampbellNessuna valutazione finora
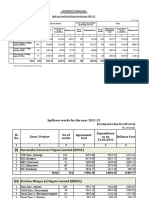
- Spill-Over and Fresh Works For The Year 2021-22: Rs. in CroresDocumento4 pagineSpill-Over and Fresh Works For The Year 2021-22: Rs. in Croreshoney honeyNessuna valutazione finora
- Engineering Hydrology Solution Manual 3rd Edition K SubramanyaDocumento162 pagineEngineering Hydrology Solution Manual 3rd Edition K Subramanyaramasamy94% (18)
- Chilton CoburnDocumento5 pagineChilton Coburnandreluisalberton100% (1)
- PPT On Sewer DesignDocumento16 paginePPT On Sewer DesignTekle GermariamNessuna valutazione finora
- Assessment of Heavy Metals (ZN, Cu, PB, CR, CD and Ni) Concentration in The Leaves, Stems and Tubers of Manihot Esculenta (Cassava) Harvested From The Egi Community, Rivers StateDocumento8 pagineAssessment of Heavy Metals (ZN, Cu, PB, CR, CD and Ni) Concentration in The Leaves, Stems and Tubers of Manihot Esculenta (Cassava) Harvested From The Egi Community, Rivers StateIjsrnet EditorialNessuna valutazione finora
- Chemical MonitoringDocumento1 paginaChemical MonitoringShamsuddin SoomroNessuna valutazione finora
- Alok NandanDocumento41 pagineAlok NandanSean Curnow100% (1)
- 428434-Offshore Drilling Rig TypesDocumento7 pagine428434-Offshore Drilling Rig TypesNagendra Singh Ranawat100% (1)
- Unit10 DeforestationDocumento25 pagineUnit10 DeforestationathulNessuna valutazione finora